Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
ЭЛЕКТРОФОРЕЗ ФРАГМЕНТОВ ДНК

2
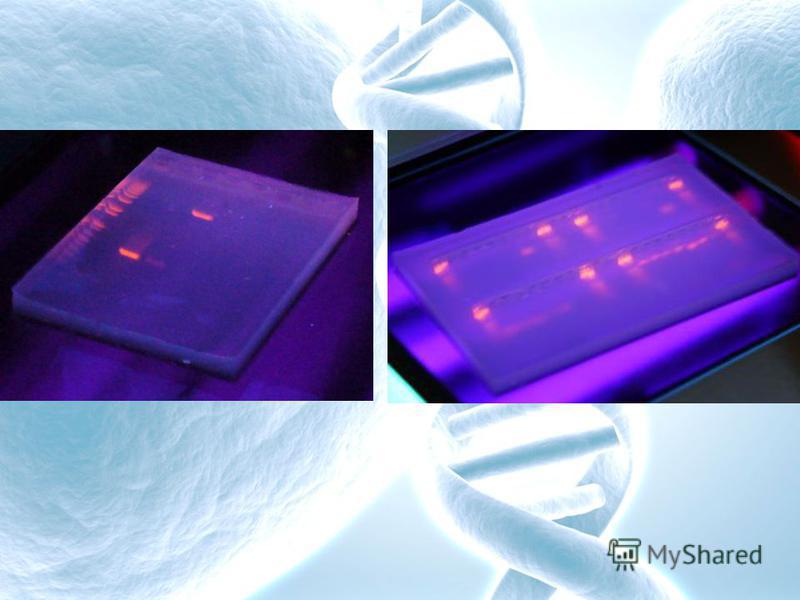
Мост из фильтровальной бумаги Агарозный гель Лунки для образцов ДНК Электрод Буферный раствор Направление движения ДНК Источник тока Рис. 1. Схематическое изображение камеры для электрофореза ДНК в агарозном геле. Справа представлена электрофореграмма фрагментов ДНК, окрашенная этидиум бромидом и помещенная под УФ свет. (М – маркеры, 1,2,3 – образцы одной и той же ДНК, разрезанные различными рестриктазами). М 1 2 3Kb

3
1. При помощи набора ферментов рестрикции исследователи научились получать фрагменты ДНК разных размеров практически из любых видов. В ходе манипуляций с различными фрагментами ДНК часто необходимо определить размер или выделить конкретный участок ДНК из смеси. Оказалось, что фрагменты ДНК легче всего разделять с помощью метода электрофореза в агарозном геле. ДНК, обработанную одной или несколькими рестриктазами, помещают в лунки застывшего агарозного геля, который помещается в специальную камеру для электрофореза (рис. 1). В камере создается электрическое поле, под действием которого фрагменты ДНК начинают перемещаться в пористом, похожем на мармелад геле. Скорость продвижения фрагментов ДНК в геле зависит от их длины. Короткие фрагменты движутся быстрее, чем длинные. Это позволяет цепочкам ДНК разной длины отделиться друг от друга. При этом фрагменты ДНК не повреждаются и их можно выделить из геля без всяких повреждений и потери биологических свойств.

4
Если после электрофореза окрасить гель специальным красителем этидиум бромидом, связывающимся с ДНК и поместить гель под ультрафиолетовый свет, то на нем будут хорошо видны окрашенные в красный цвет, расположенные на различном расстоянии друг от друга светящиеся фракции ДНК. Каждая такая фракция соответствует одному фрагменту ДНК. Следует подчеркнуть, что разные рестриктазы дают разную картину расщепления одной и той же ДНК (электрофореграмма на рис. 1 справа). Таким образом, электрофорез в агарозном геле позволяет разделить, а затем легко извлечь любые рестрикционные фрагменты ДНК в чистом виде, для последующего использования. Кроме того, анализируя электрофоретические спектры ДНК на геле, наблюдая за исчезновением одних и появлением других фракций под действием разных рестриктаз, исследователи начали составлять генетические рестрикционные карты расположения участков ДНК для разных видов.

8
ПЦР-АНАЛИЗ И СЕКВЕНИРОВАНИЕ ДНК

9
Амплификация фрагментов ДНК с помощью метода ПЦР (полимеразной цепной реакции) Несмотря на то, что методы исследования наследственного материала (нуклеиновых кислот) все время совершенствовались, тем не менее для анализа структуры ДНК требовалось определенное количество клеточного материала. Например, даже при использовании такого чувствительного метода, как фингерпринт, требуется наличие капли крови или другого эквивалентного количества образца животной или растительной ткани, содержащих в клетках достаточное для анализа количество копий ДНК.
 Несмотря на то, что методы исследования наследственного материала (нуклеиновых кислот) все время совершенствовались, тем не менее для анализа структуры ДНК требовалось опр")
10
Ситуация радикально изменилась благодаря появлению метода, который был разработан Кэри Мюллисом. Этот метод получил название полимеразной цепной реакции (ПЦР) и стал неотъемлемой процедурой, освоенной во всех генно-инженерных лабораториях мира. Использование ПЦР методики позволяет амплифицировать (размножать) ДНК или её фрагмент in vitro увеличивая количество копий в миллионы раз за несколько часов. ПЦР осуществляют в пробирке с помощью специального термостабильного фермента ДНК-полимераза (Tag-полимеразы), набора всех четырех нуклеотидов А, Т, Г и Ц и коротких олиго- нуклеотидных затравок – праймеров. Праймеры – это короткие, длиной в нуклеотидов, одноцепочечные фрагменты ДНК, комплементарные 3-концевым последовательностям копирует- мой ДНК-матрицы. Благодаря праймерам ограничивается фрагмент ДНК, который будет скопирован Tag-ДНК-полимеразой, просо- уединяющейся к 3-концам праймеров и достраивающие их до заданной длины. Полимеразная цепная реакция (ПЦР) протекает в три стадии (рис).
 и стал неотъемлемой процедурой, освоенной во всех генно-инженерных лабораториях мира. Испол")
11
Рис. 1. Последовательные стадии одного цикла амплификации (размножения) фрагмента ДНК с помощью метода полимеразной цепной реакции (ПЦР). Денатурация Гибридизация Полимеризация o С 50 o С 70 o С 5 3 t=15c. t=30c. t=90c.
 фрагмента ДНК с помощью метода полимеразной цепной реакции (ПЦР). Денатурация Гибридизация Полимеризация 3 33 3 33 3 3 3 3 3 5555 5 5 5 5 55 5 90 o С 50 o С 70 o С 5 3 t=15c. t=3")
12
1. Денатурация. Инкубационную смесь, в которой содержится образец нужной ДНК, нагревают до температуры 90 С. При этом, в течении 15 секунд происходит разрушение слабых водородных связей между нитями ДНК, и из одной двухцепочечной молекулы образуется две одноцепочечные. 2. Гибридизация праймеров. Температуру снижают до 50 С. При этом происходит гибридизация цепей ДНК с праймерами. Эта стадия обычно протекает Полимеризация. Инкубационную смесь нагревают до температуры 70 С. При этой температуре Tag-полимераза удлиняет оба праймера с их 3-концов. Праймеры дорастают до размеров матрицы. Этот процесс протекает в течении 90 секунд. В результате количество ДНК удваивается. Фер- мент Tag-полимераза была выделена из термофильных бактерий Thermus aquaticus, и отличается устойчивостью к высокой температуре. При температуре 70 С гибрид праймер-ДНК не денатурирует, а Tag-полимераза способна работать с большой скоростью. За 20 циклов амплификации количество копий ДНК достигает величины 106.

13
В последние годы удалось создать специальный прибор -- амплификатор, с помощью которого все три стадии размножения ДНК производятся автоматически, что превратило процесс ПЦР-амплификации конкретной последовательности ДНК в простую задачу. За разработку метода полимеразной цепной реакции (ПЦР) в 1993 г. Кэри Мюллис (K. Mullis) был удостоен звания лауреата Нобелевской премии.

14
Секвенирование нуклеотидных последовательностей ДНК Настоящим успехом стала разработка методов секвенирования фрагментов ДНК длиной нуклеотидных пар. Первый прямой метод определения последовательности ДНК был предложен Ф. Сэнгером в 1975 г. Он основан на элонгации ДНК при помощи фермента ДНК-полимеразы. Этим способом бы-ла быстро секвенирование короткая ДНК фага х 174, длиной 5,4 кб. Столь же мощный метод сиквенса ДНК был разработан А. Максамом и У. Гилбертом в 1977 г. в Гарвардском университете. С его помощью менее чем за год удалось установить последователь-ность ДНК для вируса sv-40 (5,2 кб) и плазмиды рBR322 (4,3 кб). Метод секвенирования ДНК по Максамому-Гилберту заключается в следующем. Один из концов фрагмента ДНК, последовательность которого нужно прочитать (секвенировать) метят с помощью 32Р. Препарат меченой ДНК делят на четыре порции и каждую из них обрабатывают реагентом, специфически разрушающим одно из четырех оснований ДНК.

15
Радиоактивная метка Двухцепочечная ДНК Цепи разделяют и получают препарат одной из них Химическим путем разрушают одно из четырех оснований, в результате чего происходит расщепление цепи в соответствующих точках. Условия реакции подбирают таким образом, чтобы в каждой точке расщеплялись только некоторые из цепей; при этом получается набор фрагментов разной длины Радиоавтограф геля Т А Г Ц Последовательность С помощью электрофореза в геле фрагменты разделяются по размеру; те из них. которые содержат радиоактивную метку, оставляют «отпечатки» на рентгеновской пленке. По положению этих отпечатков можно определить, какое именно основание было разрушено при образовании каждого из радиоактивных фрагментов ТАГЦТАГЦ Одноцепочечная ДНК ТТАГАЦЦЦГАТААГЦЦЦГЦАТТАГАЦЦЦГАТААГЦЦЦГЦА Длинные фрагменты Короткие фрагменты

16
Условия реакции подбирают таким образом, чтобы на каждую молекулу ДНК приходилось лишь несколько повреждений. Когда эти повреждённые молекулы обрабатывают пиперидином, в ДНК образуется разрыв в том месте, где находилось разрушенное основание. В результате получается набор меченых фрагментов, длины которых определяются расстоянием от разрушенного основания до конца молекулы. Например, если остатки Г находятся на расстоянии 2, 6, 11 и 16 нуклеотидов от меченого конца, как в случае рассмотренном на рисунке слева, то обработка данной цепи ДНК реагентами, разрушающими Г, приведёт к образованию меченых фрагментов длиной 2, 6, 11 и 16 нуклеотидов (при этом, естественно образуют- ся еще и фрагменты длинной 3 и 4 нуклеотида, но эти фрагменты расположенные между остатками Г будут не меченными). Наборы меченых фрагментов, образующихся при каждой из четырёх реакций подвергают электрофорезу в соседних дорожках полиакриламид него геля, при этом происходит разделение фрагментов ДНК в соответствии с их размерами. Затем проводят радио- автографию геля. Набор полос, регистрируемых на рентгеновской плёнке, «читают», определяя таким образом нуклеотидную последовательность ДНК. Так в примере представленном на рисунке последовательность цепочки ДНК от меченого конца будет следующая АЦГЦЦЦГААТАГЦЦЦАГАТТ.

17
Для анализа последовательностей ДНК на протяжении многих лет широко используется ферментативный метод, разработанный Фрэдом Сэнгером. Этот мощный метод носит название «терминация цепи» и основан на использовании дидезоксинуклеотидов (рис. 1 а). Суть метода заключается в следующем. К одноцепочечной ДНК 3'-5', которая служит матрицей, добавляется комплементарный радиоактивно меченный праймер 5'-3' и ДНК-полимераза, которая будет синтезировать комплементарную цепь 5'-3', последовательно просоединяя нуклеотиды к 3'-ОН группе предыдущих. На первом этапе набор реагентов, содержащий ДНК-матрицу 3'-5' и праймер, а также ДНК-полимеразу и набор нуклеотидов АТФ, ГТФ, ЦТФ и ТТФ, делится на 4 части и распределяется по пробиркам (рис. 1 б). Кроме того, в пробирки добавляются в небольшом количестве еще и дидезоксинуклеотиды. Причем, в каждую пробирку добавляется только один из четырех.
. Суть")
18
рис. 1 а) и дезоксирибоза; б) набор реагентов для синтеза ДНК в пробирках; в) синтез ДНК в пробирке с добавлением ддТ; г) электрофорез фрагментов ДНК.
 и дезоксирибоза; б) набор реагентов для синтеза ДНК в пробирках; в) синтез ДНК в пробирке с добавлением ддТ; г) электрофорез фрагментов ДНК.")
19
На рис. 1 в представлен ход реакции синтеза молекулы ДНК в пробирке с добавлением дидезокситимина (ддТ). Синтез новой молекулы ДНК будет останавливаться там, где в цепочку встроится дидезокситимин вместо нормального тимина, так как в дидезокситимине отсутствует группа ОН в 3' позиции. Поэтому в ряде синтезируемых ДНК репликация прервется в каждой точке, где необходим тимин (рис. 1 в). Аналогичным образом будут протекать реакции синтеза молекулы ДНК 5'-3' и в остальных трех пробирках. На втором этапе проводится электрофоретическое фракционирование синтезированных фрагментов ДНК из каждой пробирки на четырех дорожках геля. Фрагменты ДНК разделяются в соответствии с их размерами. Чем короче фрагмент ДНК, тем ближе он расположен к нижней границе геля. Анализ спектров на всех четырех дорожках геля позволяет легко установить нуклеотидную последовательность синтези- рованной 5'-3' цепочки ДНК (рис. 1 г). В нашем примере последователь- ность цепочки ДНК будет следующая 5'-ГААТЦЦГТАТГТ-3'. Соответственно последовательность комплементарной цепочки, используемой в качестве матрицы, будет 3'-ЦТТАГГЦАТАЦА-5'.
. Синтез новой молекулы ДНК будет останавливаться там, где в цепочку встроится дидезокситимин вместо нормального тимина, так как в дидезокситимине")
20
Рис. 27. Схема радиограммы сиквенса ДНК человека Г Т А Ц На рис. 27 приведена радиоавтография полиакрил- амидного геля, полученного в результате секвенирования по методу Сэнгера небольшого фрагмента ДНК важного гена человека длиной 50 нуклеотидных пар. Исходя из электрофоретического спектра ДНК представленного на радиограмме (рис. 27) можно легко определить нуклеотидную последовательность данного фрагмента уже рассмотренным нами способом. Так, просеквенированный фрагмент ДНК важного гена человека будет иметь следующую последовательность своих 50 нуклеотидов: 5'-ТАТЦАГАТЦТГЦААЦТЦА- ТАТГАТЦГАГАГГГАААТЦААТТЦТГТГААЦГ-3'. Следует отметить, что за разработку методов секвенирования ДНК Ф. Сэнгеру и У.Гилберту в 1980 году была присуждена Нобелевская премия. Интересно, что у Сэнгера это была уже вторая Нобелевская премия. Первую он получил в 1958 году по химии за открытие строения инсулина.

21
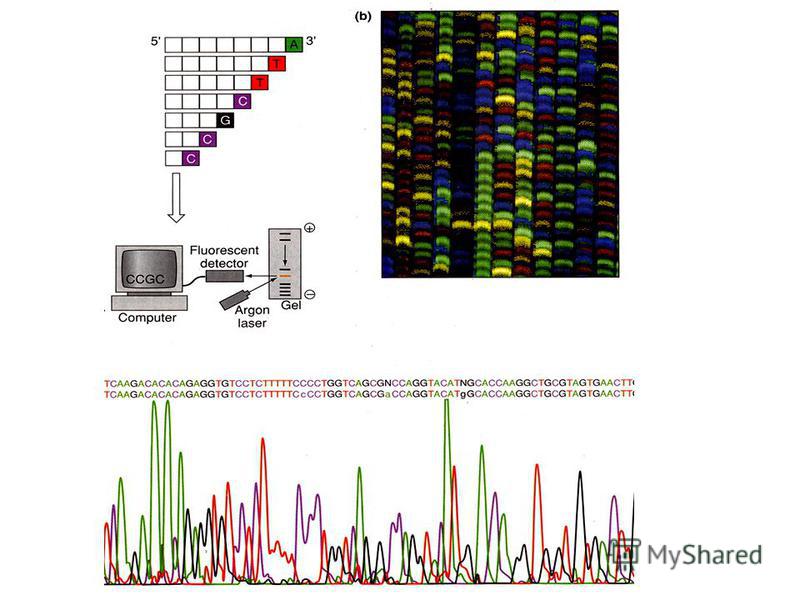
Стало ясно, что методы секвенирования нужно как-то автоматизировать. Это сделал в 1986 году Лерой Худ. Он усовершенствовал метод Сэнгера и вместо радиоактивных меток при анализе последовательностей ДНК стал использовать флюорисцентные красители разного цвета для четырех дидезоксинуклеотидов. Флюорисцентный анализ позволил все четыре смеси электрофорезировать вместе, как представлено на рис. 2 б, что дало возможность определять последовательность ДНК автоматически и выводить результат на экран компьютера (рис. 2 в).

22
Рис. 2 а) электрофорез фрагментов ДНК на четырех дорожках геля; б) электрофорез фрагментов ДНК с флюоресцентными красителями на одной дорожке; в) автоматическое прочтение последовательности нуклеотидов ДНК.
 электрофорез фрагментов ДНК на четырех дорожках геля; б) электрофорез фрагментов ДНК с флюоресцентными красителями на одной дорожке; в) автоматическое прочтение последовательности нуклеотидов ДНК.")
23
На этом принципе основана работа современных секвенаторов – приборов для автоматического анализа последовательностей ДНК. С помощью секвенаторов в настоящее время генетики могут «прочитывать» 1000 и более нуклеотидных пар за одну операцию.

Еще похожие презентации в нашем архиве:







 изобрел в 1983 году американский ученый Кэри Мюллис.")













