Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

2
Радиобиология как предмет. История радиобиологии. Радиочувствительность. Физические основы действия излучений на биологические объекты. Прямое и косвенное действие ионизирующих излучений. Молекулярные аспекты биологического действия ионизирующих излучений. Проявления лучевого поражения на уровне клетки. Факторы, определяющие выживаемость клеток после облучения. Модификация радиочувствительности. Кислородный эффект. ОБЭ ионизирующих излучений. Механизм действия ИИ. Радиочувствительность тканей, органов, организма. Детерминированные эффекты. Процессы восстановления в облученном организме. Лучевая болезнь человека. Биологическое действие инкорпорированных радиоактивных веществ. Опосредованные эффекты облучения. Отдалённые последствия облучения. Последствия облучения эмбриона и плода. Фармакохимическая противолучевая защита организма. Радиобиологические основы лечебного применения ионизирующих излучений. Источники облучения человека. Медицинские последствия радиационных аварий. Регламентация облучения человека.

3
Прямое и косвенное действие ионизирующих излучений. Молекулярные аспекты биологического действия ионизирующих излучений.

4
Прямое и косвенное действие ионизирующих излучений Прямое действие ионизирующего излучения заключается в поглощении энергии молекулой или атомом, в результате чего происходит ионизация молекул и, как следствие, изменения структуры макромолекул: белков и ДНК. Косвенное действие ионизирующего излучения заключается в образовании нескольких высокореакционных продуктов из молекулы воды (радиолиз ) и растворённых в ней низкомолекулярных соединений, а не энергий излучения, поглощённой самими исследуемыми молекулами. Мишень Излучение Н2ОН2О Н*ОН* + Мишень Прямое действие излучения Косвенное действие излучения

5
Схема первичных физико-химических процессов на пути от ионизации к конечному биологическому эффекту.. Падающий фотон или заряженная редко- ионизирующая частица Падающий быстрый нейтрон, дающий протоны отдачи, или тяжелые заряженные частицы Молекула или атом Ионные пары Свободные радикалы Химические изменения вследствие разрыва связей Биологический эффект

6
Молекулярные аспекты биологического действия ионизирующих излучений. Повреждения ДНК Задержка прохождения клеточного цикла Индукция апоптоза

7
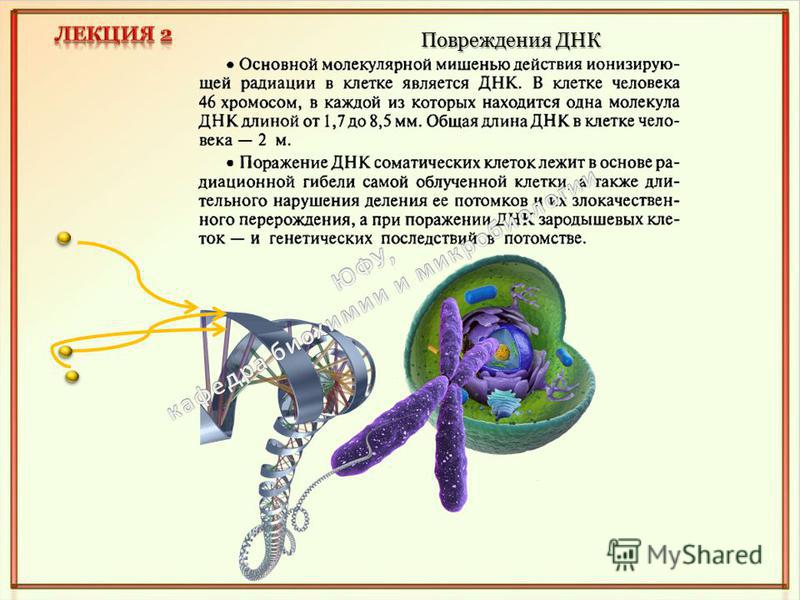
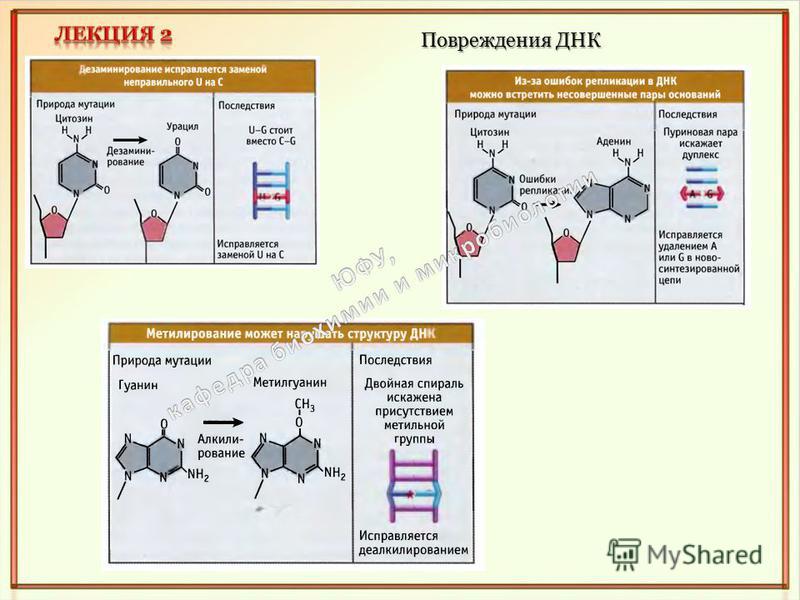
Повреждения ДНК

10
III.Метилирование IV.Размыкание кольца V. Окисление VI.Образование тимьяновых димеров

11
Повреждения ДНК

12
Задержка прохождения клеточного цикла

13
Сверочные точки

14
Задержка прохождения клеточного цикла

16
Проявление лучевого поражения на уровне клеток Хроматидные и хромосомные аберрации Микроядра Уменьшение клоногенного потенциала Апоптоз и некроз «Коммунальный эффект» Генетическая нестабильность Повышенная гибель облученных клеток

17
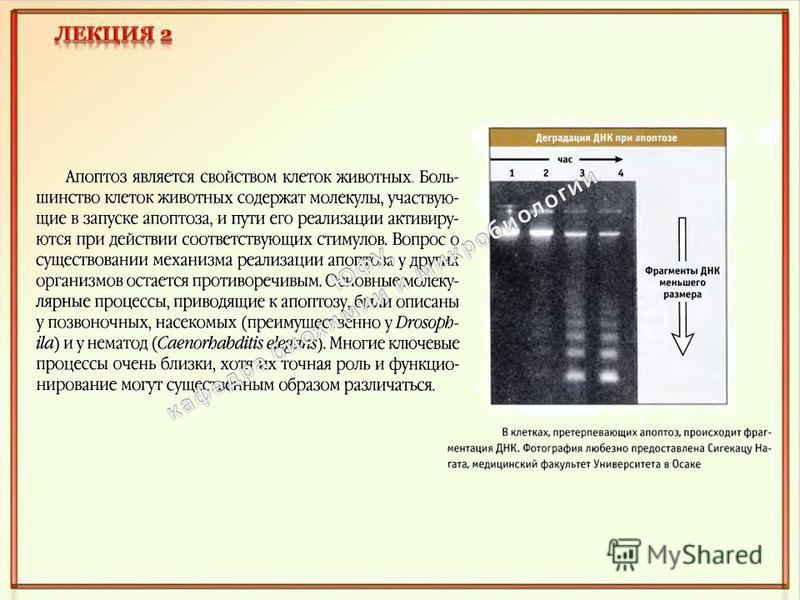
Термин"апоптоз» был предложен в 1972 г. английскими учеными J.F.R. Кеrr, А.Н. Wyllie и A.R. Currie и состоит из двух греческих слов, означающих в буквальном смысле "отделение лепестков от цветов", а применимо к клетке - особый тип смерти путем разделения ее на части с образованием апоптозных телец, которые впоследствии фагоцитируются соседними клетками разного типа.

18
Существует два варианта гибели животных клеток: апоптоз и некроз.

19
Сравнение апоптоза и некроза Апоптоз Некроз Уменьшение объёма клетки Сморщивание цитоплазматической мембраны Конденсация ядра Разрыв нити ДНК Распад ядра на части Фрагментация клетки на мембранные везикулы с внутриклеточным содержимым (апоптозные тельца) Нарушение асимметрии мембраны, появление в наружном слое фосфатидилсерина и фосфатидилэтаноламина Обнаружение апоптозных клеток макрофагами и фагоцитоз Непрограммируемая патологическая форма клеточной смерти Разрыв цитоплазматических и внутриклеточных мембран Разрушение органелл Выход лизосомальных ферментов в цитоплазму Излияние цитоплазмы в межклеточное пространство Обязательное наличие воспаления (rubor, tumor, calor, dolor, functio laesa – покраснение, опухание, жар, боль, нарушение функций)

22
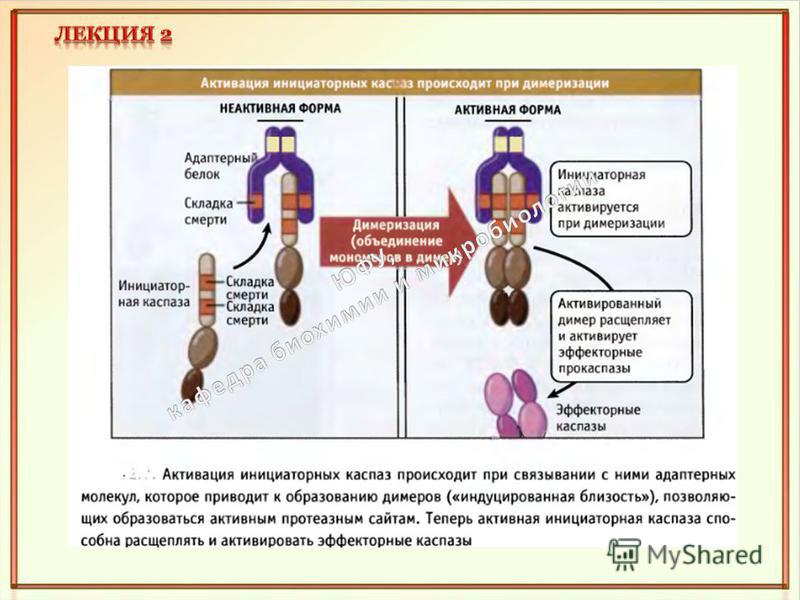
Каспазы относятся к семейству эволюционно консервативных протеаз – ферментов, катализирующих ограниченное расщепление клеточных белков. Известны 14 членов семейства: каспара-1, каспара-2, каспара-3 и т.д. В активном центре фермента – остаток цистеина. Все они являются а с пазами: специфически узнают определенные тетра пептидные звенья белков и расщепляют пептидную связь по карбоксильному концу остатка аспарагиновой кислоты. Каспаза (от англ. caspase), где буква c соответствует цистеину (cysteine), корень asp – аспартату (aspartate), ase – суффиксу в названиях ферментов. В клетке каспары синтезируются в форме латентных предшественников – проферментов, называемых прока с пазами. Структура каспары-7 с ингибитором. X X AspX X Caspase Разрезание пептидной связи по карбоксильному концу Asp.

23
Виды каспар Эффекторные каспары Каспазы участвующие в воспалении Инициаторные каспары Casp 3, Casp 6, Casp 7 Casp 2, Casp 8, Casp 9

25
Структурно прокаспара (молекулярная масса до 50 к Да) состоит из трех звеньев: 1. N-концевого звена (про домена) 2. Промежуточного домена, предшественника большой субъединицы (~ 20 к Да) 3.С-концевого домена, предшественника малой субъединицы (~ 10 к Да) зрелого фермента. Продомены инициирующих (а также воспалительных) прокаспар содержат свыше 100 аминокислотных остатков. Они выполняют важную функцию в активации фермента: осуществляют взаимодействие прокаспар с белками- адаптерами. В этих белок-белковых взаимодействиях участвуют специализированные участки про доменов, у разных про- каспар это: I. DED (death effector domain – домен эффектора смерти) II. CARD (caspase recruitment domain – домен рекрутирования каспары) III. DID (death inducing domain – домен, вызывающий смерть).
 состоит из трех звеньев: 1. N-концевого звена (про домена) 2. Промежуточного домена, предшественника большой субъединицы (~ 20 к Да) 3.С-концевого домена, предшественника малой субъединицы (~ 10 к")
27
CAD ( (caspase-activated DNA-ase )CAD I CAD (DFF) CADCAD I CAD Подвергается протеолизу ингибитор ДНКазы, ответственной за фрагментацию ДНК. В нормальных клетках апоптозная ДНКаза CAD (caspase-activated DNase – ДНКаза, активируемая каспарой) образует неактивный комплекс с ингибитором CAD, обозначаемым I CAD или DFF (DNA fragmentation factor – фактор фрагментации ДНК). При апоптозе ингибитор I CAD с участием каспар-3 или -7 инактивируется и свободная CAD, вызывая межнуклеосомальные разрывы хроматина, ведет к образованию фрагментов ДНК с молекулярной массой, кратной молекулярной массе ДНК в нуклеосомных частицах, – 180–200 пар нуклеотидов. Эти фрагменты при электрофоретическом разделении в агарозном геле дают характерную лесенку ДНК. Апоптоз возможен и без фрагментации ДНК.
CAD I CAD (DFF) CADCAD I CAD Подвергается протеолизу ингибитор ДНКазы, ответственной за фрагментацию ДНК. В нормальных клетках апоптозная ДНКаза CAD (caspase-activated DNase – ДНКаза, активируемая каспарой) образует")
28
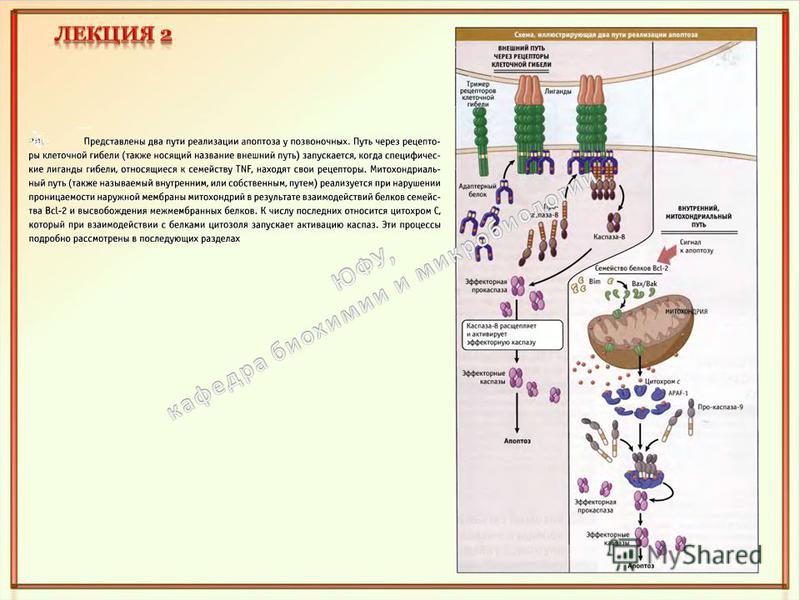
Митохондрии выполняют центральную роль в апоптозе, при этом наблюдается увеличение проницаемости митохондриальной мембраны. Баланс между про- и анти-апоптозных членов семейства Bcl-2 регулирует выход про-апоптозных веществ из митохондрий, ведущих к запуску апоптоза, таких как AIF, эндонуклеаза G, Smac/DIABLO и цитохром C. Утечка цитохрома- С из митохондрии приводит к образованию апоптосомы в цитоплазме, которая активирует каспару-9 и запускает клеточную смерть. Оба пути приводят к активации каспар и запуску каскада реакций приводящих к гибели клетки. Рецепторы апоптоза - семейства белков CD95 (Apo-1 или Fas) и TNF-R (фактор опухолевого некроза). TNF-альфа высоко цитотоксичная молекула, использовалась как лекарство против рака. TNF-R1 рецептор широко распространен и поэтому не может быть избирательным. Другие представители этого семейства (не все) имеют домен клеточной смерти (DD) - домен белок- белкового взаимодействия связывающийся с белком адаптером, таким как FADD. Активация рецепторов апоптоза лигандами (например, CD- 95L и TNF-альфа приводит к активации каспары- 8, запуская каскад реакций ведущих к апоптозу.

30
Рецепторы апоптоза - семейства белков CD95 (Apo-1 или Fas) и TNF-R (фактор опухолевого некроза). TNF-альфа высоко цитотоксичная молекула, использовалась как лекарство против рака. TNF-R1 рецептор широко распространен и поэтому не может быть избирательным. Другие представители этого семейства (не все) имеют домен клеточной смерти (DD) - домен белок- белкового взаимодействия связывающийся с белком адаптером, таким как FADD. Активация рецепторов апоптоза лигандами (например, CD- 95L и TNF-альфа приводит к активации каспары- 8, запуская каскад реакций ведущих к апоптозу. Белок TRADD опосредует передачу сигнала от активированных рецепторов TNF к белкам FADD и RIP.
 и TNF-R (фактор опухолевого некроза). TNF-альфа высоко цитотоксичная молекула, использовалась как лекарство против рака. TNF-R1 рецептор широко распространен и поэтому не может быть избирател")
32
Радиационная задержка клеточного деления Задержка в продвижении по циклу дает клетке больше времени для репарации ДНК перед S-фазой или митозом, когда целостность молекулы ДНК особенно значима для нормальной жизнедеятельности. Однако данные о более длительной задержке в наступлении митоза у погибающих впоследствии клеток по сравнению с сохраняющими клоногенный потенциал указывают, по-видимому, на более глубокое повреждение ДНК данной клетки. Снижение числа делящихся клеток после облучения было замечено уже вскоре после открытия рентгеновских лучей, что и послужило одним из оснований к их применению для подавления опухолевого роста. Задержка в наступлении очередного деления наблюдается почти у всех клеток облучаемой популяции, причем ее длительность зависит от дозы ионизирующего излучения.

33
Радиационная задержка клеточного деления

34
Хромосомные аберрации и микроядра

35
АБЕРРАЦИИ Хромосомные Хроматидные Хромосомные аберрации возникают в случае, когда клетка подверглась облучению в предсинтетической стадии цикла или в S-периоде, но до начала удвоения определенного участка своего генома. При неверном воссоединении оторванных друг от друга фрагментов ДНК такое нарушение воспроизводится во время репликации (удвоении). Итогом является образование дицентриков хромосом, имеющих две центромеры, что может сопровождаться появлением ацентрических фрагментов, хорошо видных при сравнении метафазных пластинок облученных и не облученных лимфоцитов Хроматидные аберрации возникают в клетке, облученной уже после завершения репликации всей ДНК или того ее участка, разрыв которого и приведет к формированию аберрации. Разрыв одной из хроматид проявится в виде ее укорочения и образования ацентрического фрагмента, который будет виден при мета- или анафазном анализе. Разрыв обеих хроматид может проявиться различным образом образованием двух фрагментов, их слиянием в один (принято говорить о «липкости» концов разорванных хроматид и появлением кольцевой хромомомы.

36
Хромосомные аберрации и микроядра Дицентрик Две центромеры Разрыв хромосом Удаляемый генетический материал Слияние Удаляемый генетически й материал Разрыв хромосом Делеция

37
Хромосомные аберрации и микроядра Парацентрическая инверсия Перицентрическая инверсия Инверсия Разрыв хромосом Центромера Инверсия Инверсия ДНК

38
Хромосомные аберрации и микроядра Кольцевая хромосома Разрыв хромосом Слияние Удаляемый генетический материал

39
Хромосомные аберрации и микроядра Сбалансированная транслокация Несбалансированна я транслокация Хромосома А Хромосома В Нормальная хромосома 1 родителя Хромосома от родителя 2 с сбалансированной транслокацией

40
Хромосомные аберрации и микроядра

41
Микроядра

42
Утеря клоногенного потенциала и кривые выживания Подсчет хромосомных аберраций или микроядер позволяет довольно точно оценить радиочувствительность клеток, однако оба метода весьма трудоемки. Основные данные о радиочувствительности получены путем изучения выживаемости клеток, с помощью оценки их клоногенной способности, под которой понимается способность клетки образовывать видимую невооруженным глазом колонию. Согласно договоренности исследователей, клетка после облучения считается «выжившей», если она образует колонию, состоящую из более чем 50 клеток. Учет выживаемости проводится в сроки, достаточные для образования крупных колоний всеми интактными клетками. Для образования видимой глазом колонии облученная клетка должна совершить не менее 5 полностью успешных делений, т. е. делений, в результате которых дочерние клетки также будут способны к делению.

43
Кривые выживаемости в области малых доз В последнее десятилетие большое внимание исследователей привлечено к изучению формы кривой дозаэффект в области малых (до 0,8 Гр) доз излучения. Этот диапазон наиболее интересен с позиций радиационной безопасности, так как при профессиональном контакте с излучениями, а также при радиационных авариях наибольшее число лиц подвергается воздействию именно в этих дозах. При лучевой терапии фракционированному воздействию в таких дозах подвергаются участки нормальных тканей на краях поля облучения, где полученная доза спадает с классических 2 Гр за фракцию до нуля. Обнаружено, что кривая дозаэффект у ряда радиоустойчивых линий клеток на начальном участке, в зоне «плеча», с ростом дозы снижается не плавно, а вначале очень быстро, затем следует плато или даже наблюдается небольшой подъем выживаемости, после чего она принимает обычный вид.
 доз излучения. Этот диапазон наиболее интересен с позиций радиационной безопасно")
44
«Коммунальный» эффект Коммунальный эффект, также называемый «эффектом свидетеля» (по англ. bystander effect), заключается в поражении клеток, находящихся вне зоны воздействия радиации, но контактирующих тем или иным способом с облучаемыми клетками. При этом имеется в виду как непосредственный контакт облученных и не облученных клеток, так и их нахождение в одном и том же культуральном сосуде, а в ряде случаев и контакт необлученных клеток со средой, в которой другие клетки подвергались облучению. Изучение образования соединений, передающих сигнал об облучении на интактные клетки, и каскада биохимических реакций, возникающих в клетках в ответ на поступление такого сигнала, указывает на участие в этом процессе такого универсального передатчика сигналов, как окись азота NО. При исследовании роли непосредственного межклеточного взаимодействия показано участие в осуществлении коммунального эффекта белка коннексина 43 межклеточного передатчика сигналов. Введение антител к фактору некроза опухолей а в среду также подавляет выраженность коммунального эффекта. Однако в целом механизм этого Удивительного явления остается малоизученным.
, заключается в поражении клеток, находящихся вне зоны воздействия радиации, но контактирующих тем или иным способом с облучаемыми клетками. П")
Еще похожие презентации в нашем архиве:






 Глубокие нарушения жизнедеятельности вызываются.")


















