Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
ВОЕННО–МЕДИЦИНСКАЯ АКАДЕМИЯ имени С.М. Кирова Кафедра биологической и медицинской физики ЛЕКЦИЯ 14 по дисциплине «Физика, математика» на тему: «Распространение возбуждения по возбудимым мембранам. Биофизические основы электрокардиографии» для курсантов и студентов I курса ФПВ, ФПиУГВ, спецфакультета

2
1. Распространение потенциала действия по возбудимым мембранам При возбуждении нервного волокна (например, прямоугольным импульсом напряжения) можно зарегистрировать потенциалы действия не только в месте раздражения, но и на значительных расстояниях от него.
 можно зарегистрировать потенциалы действия не только в месте раздражения, но и на значительных расстояниях")
3
На всем протяжении нервного волокна ПД имеют одинаковую амплитуду, но появляются с задержкой, которая пропорциональна расстоянию от места нанесения стимула. Например, в двигательном нерве ПД регистрируется на участке, расположенном от места раздражения на расстоянии 1 м, через 10 мс; отсюда следует, что скорость распространения возбуждения по нерву равна 100 м/с.

4
Распространение возбуждения по нерву складывается из двух последовательных процессов: 1) распространения ЭМП с затуханием (декрементом); 2) ретрансляции ПД.
 распространения ЭМП с затуханием (декрементом); 2) ретрансляции ПД.")
5
Пусть в некоторой точке нервного волокна (аксона) развился и достиг пика ПД, т.е. произошла деполяризация мембраны. В месте возникновения ПД потенциал внутренней стороны мембраны положителен, а потенциал наружной стороны мембраны отрицателен.
 развился и достиг пика ПД, т.е. произошла деполяризация мембраны. В месте возникновения ПД потенциал внутренней стороны мембраны положителен, а потенциал наружной стороны мембраны отрицателен.")
6
И в цитоплазме, и в окружающей мембрану межклеточной жидкости возникают ионные токи (локальные токи): между участками поверхности мембраны с большим потенциалом (положительно заряженными) и участками поверхности мембраны с меньшим потенциалом (отрицательно заряженными).
: между участками поверхности мембраны с большим потенциалом (положительно заряженными) и участками поверхности мембраны с меньшим потенциалом (отрица")
8
За счет этих токов потенциал внутренней поверхности соседних невозбужденных участков мембраны повышается (становится более положительным), потенциал наружной поверхности невозбужденных участков понижается (становится более отрицательным).
, потенциал наружной поверхности невозбужденных участков понижается (становится более отрицательным).")
9
Трансмембранная разность потенциалов уменьшается по абсолютной величине, невозбужденные участки мембраны деполяризуются.

10
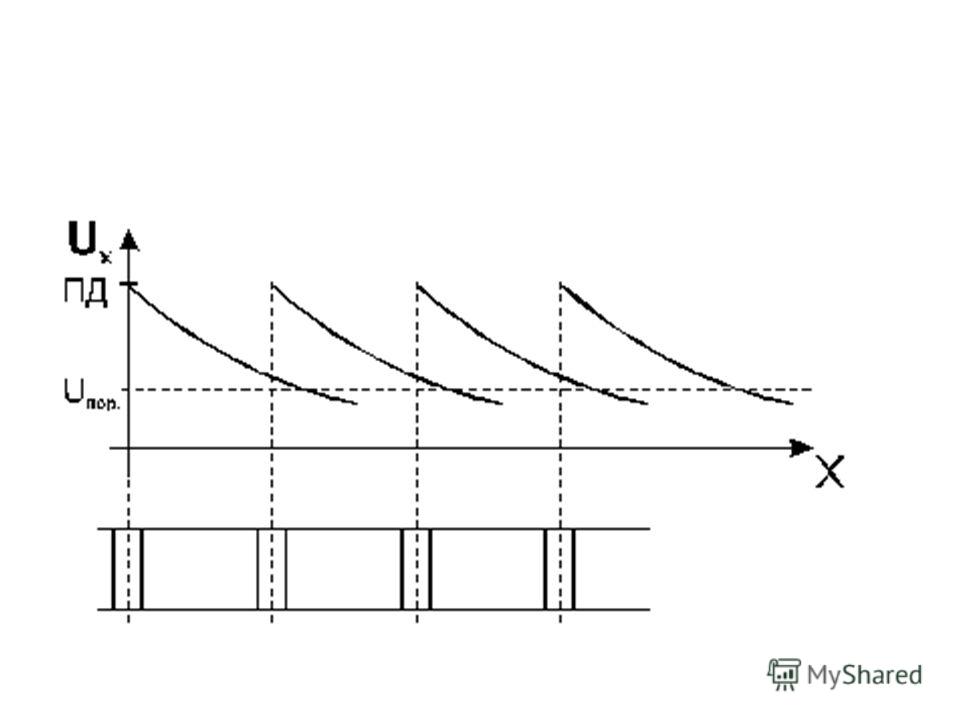
По мере удаления от точки возникновения ПД изменения трансмембранной разности потенциалов убывают по экспоненциальному закону (распространение с затуханием или декрементом).
.")
11
В тех точках мембраны, где сдвиг трансмембранной разности потенциалов оказывается выше КМП, открываются натриевые каналы и происходит развитие новых ПД (ретрансляция потенциала действия).
.")
12
Таким образом, возникающий на возбудимой мембране ПД является надпороговым стимулом для определенного участка мембраны.

13
Поскольку распространение ЭМП происходит со скоростью света в среде, пассивные сдвиги трансмембранной разности потенциалов происходят быстро, и скорость распространения возбуждения по мембране зависит от величины участка, одновременно охваченного возбуждением.

14
Величина деполяризующего потенциала зависит от расстояния от возбужденного участка мембраны следующим образом: U x = U 0. e -x/λ

15
где U x – величина деполяризующего потенциала в точке "х"; U 0 – изменение мембранного потенциала в точке возбуждения; х – расстояние от места возникновения возбуждения; λ – постоянная длины мембраны (равная расстоянию, на котором деполяризующий потенциал уменьшается в "е" раз).

16
Постоянная длины определяется следующими параметрами нервного волокна:

17
где r m – удельное электрическое сопротивление оболочки волокна; δ – толщина оболочки; а – радиус волокна; r i – удельное сопротивление цитоплазмы.

18
Чем больше константа длины мембраны, тем меньше затухание и выше скорость распространения нервного импульса. Величина λ тем больше, чем больше радиус аксона и удельное сопротивление мембраны и чем меньше удельное сопротивление цитоплазмы.

19
Большую скорость распространения нервного импульса по аксону кальмара обеспечивает их гигантский по сравнению с аксонами позвоночных диаметр, равный 1- 2 мм (λ ~ 2,5 мм).
.")
20
Такой способ повышения скорости распространения возбуждения посредством утолщения нервных волокон пригоден для животных, у которых мало быстропроводящих коммуникаций.

21
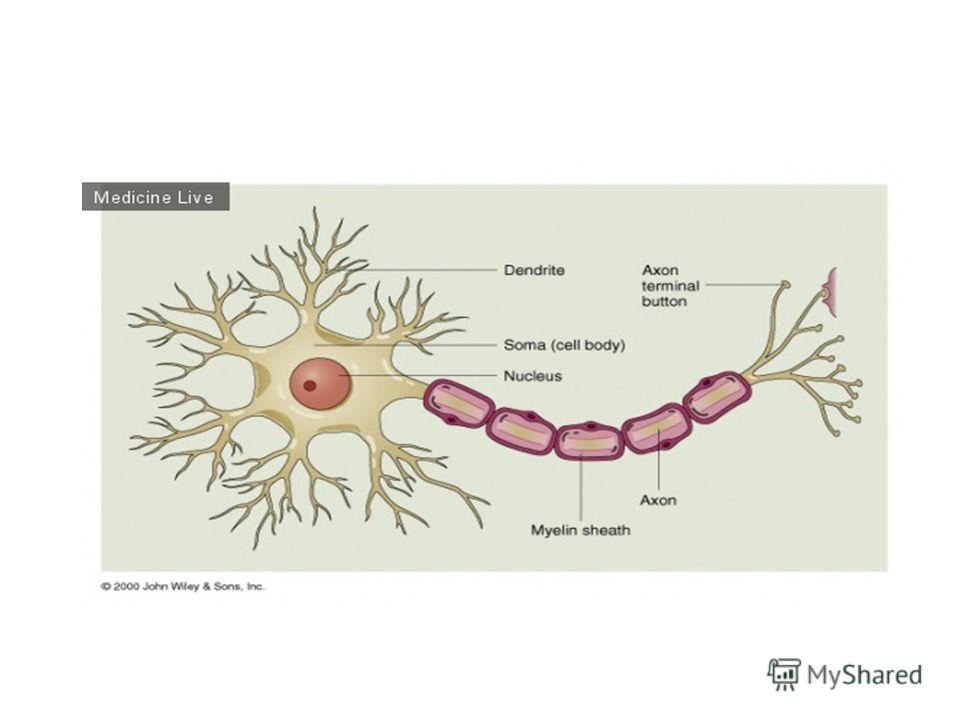
У позвоночных животных, которые имеют нервы с большим количеством проводящих волокон, возможности их утолщения ограничены размерами животного. Большая скорость передачи возбуждения в нервных волокнах достигается другими способами. Аксоны позвоночных снабжены миелиновой оболочкой.

24
Миелиновая оболочка образуется в процессе наматывания на аксон окружающих его шванновских клеток. Оболочка представляет собой многомембранную систему, включающую несколько десятков элементарных клеточных мембран, прилегающих друг к другу.

25
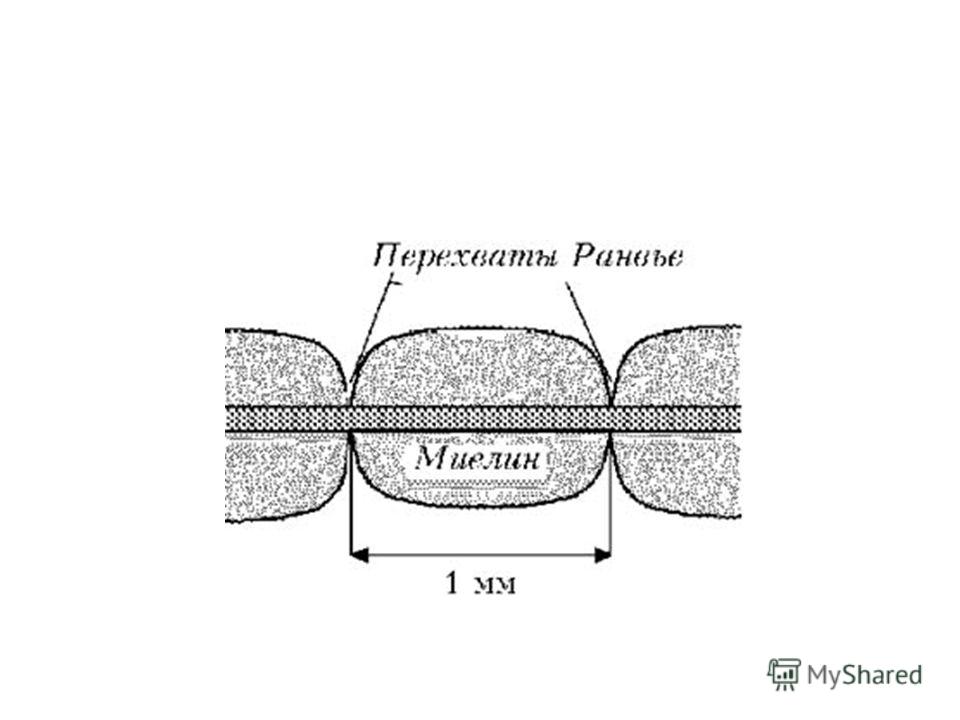
Диффузия ионов через миелиновую оболочку невозможна. Поэтому в мякотном волокне генерация ПД возможна только там, где миелиновая оболочка отсутствует (в перехватах Ранвье или активных узлах). В среднем расстояние между перехватами Ранвье составляет около 1 мм.
. В среднем расстояние между перехватами Ранвье составляет о")
26
Мембрана перехвата Ранвье специализирована для генерации возбуждения: плотность натриевых потенциалзависимых каналов здесь примерно в 100 раз выше, чем в немиелинизированных нервных волокнах.

27
От перехвата к перехвату возбуждение распространяется за счет декрементного распространения ЭМП.

29
При этом постоянная длины (λ) для этих волокон больше (так как увеличиваются сопротивление мембраны и ее толщина). Высокое значение постоянной длины обеспечивает высокую скорость распространения возбуждения по мякотным волокнам (до 140 м/с).
 для этих волокон больше (так как увеличиваются сопротивление мембраны и ее толщина). Высокое значение постоянной длины обеспечивает высокую скорость распространения возбуждения по мякотным волокнам (до 140 м/с).")
30
Ретрансляция ПД обычно происходит на 2-3 соседних перехватах Ранвье. Более частое, чем необходимо для обеспечения нормального распространения возбуждения, расположение активных узлов служит повышению надежности нервных коммуникаций в организме.

31
Поскольку ретрансляция ПД происходит только в перехватах Ранвье, то возбуждение как бы "перепрыгивает" через миелинизированные участки мембраны; такой тип проведения возбуждения получил название сальтаторного (saltus (лат.) = скачок).
 = скачок).")
32
Миелинизация обеспечивает повышение скорости проведения при существенной экономии энергетических ресурсов. Потребление кислорода такими волокнами в 200 раз меньше, чем при непрерывном распространении нервных импульсов по безмякотным аксонам

33
2. Синаптическая передача. Функциональный межклеточный контакт, обеспечивающий переход возбуждения с одной клетки на другую, получил название сИнапса (от греч. глагола "синапто" – смыкать).
.")
34
Существует два принципиально различных типа синапсов – электрические и химические.

35
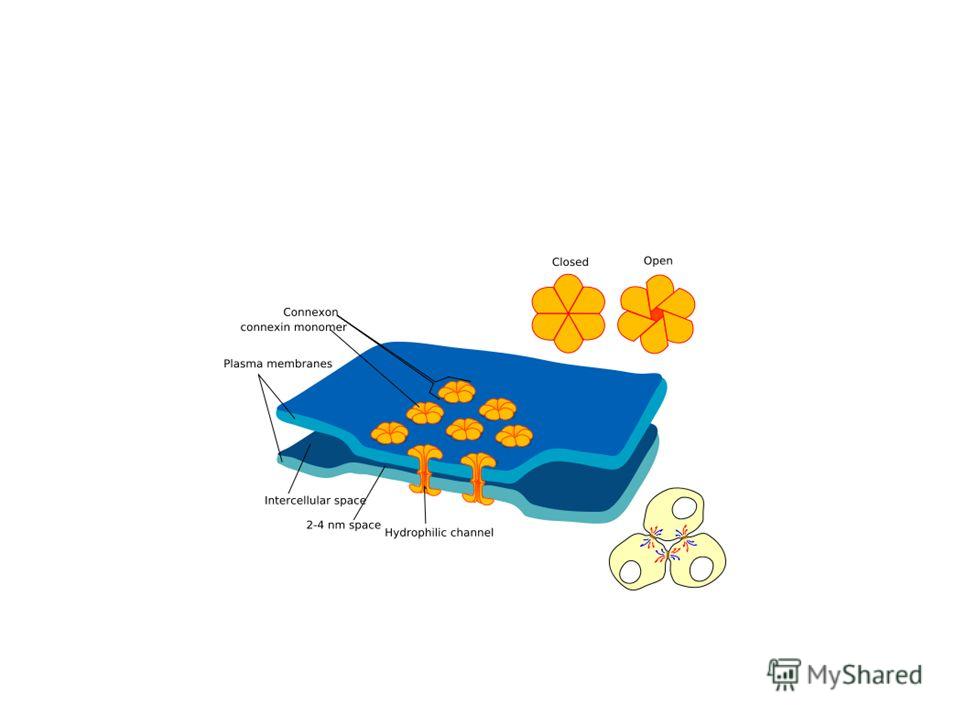
Электрическая синаптическая передача возможна только при очень тесном соприкосновении взаимодействующих клеток – при расстоянии между ними не более нанометров (часто 2-4 нм).
.")
36
В этом случае развитие ПД на мембране одной клетки приводит за счет возникновения локальных токов к деполяризации мембраны другой клетки, которая может оказаться выше порога генерирования ПД.

38
Большое значение для осуществления электрической передачи нервного импульса имеет существование в области синапса особых межклеточных контактов – щелевых контактов (нексусов). При этом в каждой из двух соседних мембран находятся регулярно расположенные коннексоны (канальные белки с большим диаметром канала и, соответственно, высокой проводимостью для ионов, и даже более крупных молекул с молекулярной массой до 1000).
. При этом в каждой из двух соседних мембран находятся регулярно расположенн")
40
Такие контакты обычны для ЦНС, миокарда и гладкой мускулатуры, где связанные щелевыми контактами клетки образуют функциональный синцитий (возбуждение переходит от одной клетки к другой очень быстро и без заметного снижения амплитуды потенциала действия на границе).

41
Щелевые контакты регулируемы, они могут закрываться при снижении рН или повышении концентрации Са 2+ (повреждение клеток или глубокие нарушения обмена). За счет такого механизма пораженные места изолируются от остальной части синцития, и распространение патологии ограничивается (инфаркт миокарда).
. За счет такого механизма пораженные места изолируются от остальной части синцития, и распростране")
42
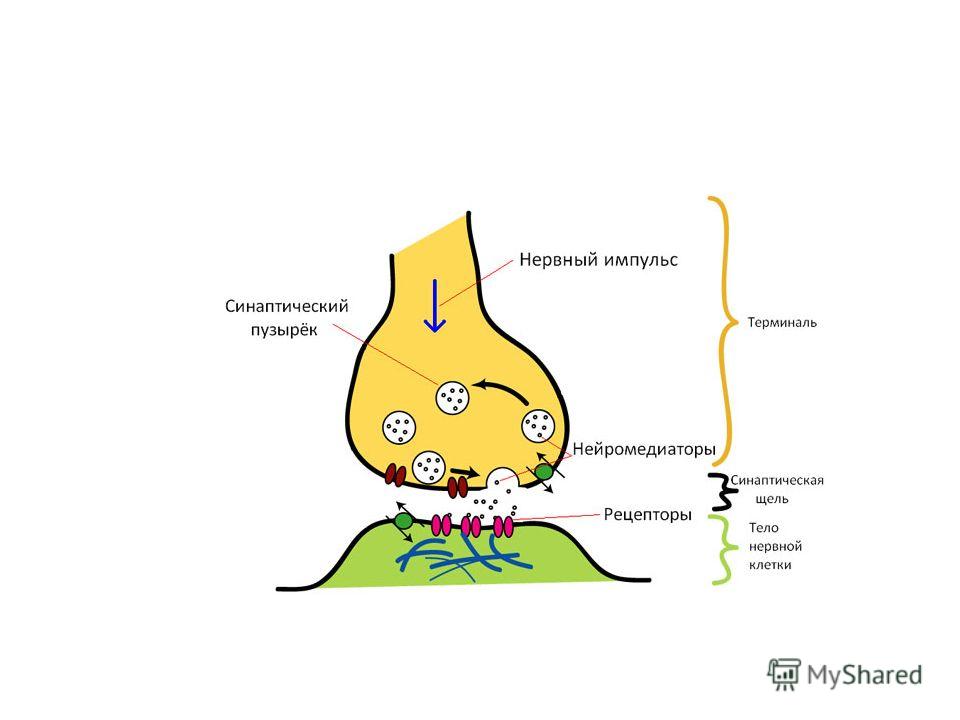
Химическая синаптическая передача осуществляется с помощью химических веществ-посредников (медиаторов). В этом случае расстояние между взаимодействующими клетками в области контакта (ширина синаптической щели) больше.
. В этом случае расстояние между взаимодействующими клетками в области контакта (ширина синаптической щели) больше.")
43
Электрическое поле затухает в пределах синаптической щели и не может деполяризовать постсинаптическую мембрану. Отсюда возникает необходимость химического посредника.

45
Деполяризация пресинапса приводит к изменению проницаемости пресинаптической мембраны для медиатора, медиатор выбрасывается в синаптическую щель, диффундирует через нее и взаимодействует с белками- рецепторами постсинаптической мембраны.

46
Изменение конформации белков- рецепторов при образовании комплекса "рецептор-медиатор" приводит к открытию на мембране специфических химиочувствительных ионных каналов, протекающие через которые ионные токи изменяют мембранный потенциал на мембране.

47
В зависимости от направления изменения трансмембранного потенциала химические синапсы могут быть возбуждающими (деполяризация постсинаптической мембраны) или тормозными (гиперполяризация постсинаптической мембраны).
 или тормозными (гиперполяризация постсинаптической мембраны).")
48
В случае возникновения ВПСП (возбуждающего постсинаптического потенциала) он с затуханием (декрементом) распространяется по постсинаптической мембране и может вызвать возникновение ПД на возбудимых участках мембраны принимающей сигнал клетки, если он превышает пороговый уровень.
 он с затуханием (декрементом) распространяется по постсинаптической мембране и может вызвать возникновение ПД на возбудимых участках мембраны принимающей сигнал клетки, если он")
49
3. Особенности биоэлектрогенеза мышечных волокон миокарда сердца. Сердце выполняет в кровеносной системе роль четырехкамерного насоса, обеспечивающего движение крови по сосудам. Оно представляет собой полый мышечный орган, состоящий из четырех отделов – двух предсердий и двух желудочков.

50
Ритмические сокращения сердца возникают под действием импульсов (ПД), зарождающихся в нем самом. Если изолированное сердце поместить в соответствующие условия, то оно будет продолжать биться с постоянной частотой. Это свойство называется автоматизмом.
, зарождающихся в нем самом. Если изолированное сердце поместить в соответствующие условия, то оно будет продолжать биться с постоянной частотой. Это свойство называется автоматизмом")
51
Функциональным элементом сердца служит мышечное волокно – цепочка из клеток миокарда, соединенных "конец в конец" и заключенных в общую саркоплазматическую оболочку (основную мембрану).
.")
52
В зависимости от морфологических и функциональных особенностей различают два типа волокон миокарда:

53
1) волокна рабочего миокарда предсердий и желудочков, составляющие его основную массу и обеспечивающие нагнетательную функцию (типичные миокардиальные волокна = ТМВ);
 волокна рабочего миокарда предсердий и желудочков, составляющие его основную массу и обеспечивающие нагнетательную функцию (типичные миокардиальные волокна = ТМВ);")
54
2) волокна водителя ритма (пейсмекера) и проводящей системы (атипичные мышечные волокна), отвечающие за генерацию возбуждения и проведение его к клеткам рабочего миокарда.
 волокна водителя ритма (пейсмекера) и проводящей системы (атипичные мышечные волокна), отвечающие за генерацию возбуждения и проведение его к клеткам рабочего миокарда.")
55
Миокард (сердечная мышца), подобно нервным тканям и скелетным мышцам, принадлежит к возбудимым тканям. Это значит, что клетки миокарда обладают потенциалом покоя (ПП), отвечают на надпороговые стимулы генерацией потенциала действия (ПД) и способны проводить ПД без затухания (бездекрементно).
, подобно нервным тканям и скелетным мышцам, принадлежит к возбудимым тканям. Это значит, что клетки миокарда обладают потенциалом покоя (ПП), отвечают на надпороговые стимулы генерацией потенциала действия (ПД) и способны пр")
56
Межклеточные соединения (щелевые контакты) способствуют проведению возбуждения и обеспечивают функционирование миокарда как функционального синцития (т.е. возбуждение, возникшее в каком-либо из отделов сердца, охватывает все без исключения невозбужденные волокна).
 способствуют проведению возбуждения и обеспечивают функционирование миокарда как функционального синцития (т.е. возбуждение, возникшее в каком-либо из отделов сердца, охватывает все без исключения невозбужде")
57
Как и в нервных клетках и волокнах скелетных мышц, ПД в типичных миокардиальных волокнах возникает в ответ на стимул (переданный с АТМВ ПД) и начинается с быстрой реверсии мембранного потенциала от ПП (примерно - 90 мВ) до потенциала инверсии (примерно + 30 мВ).
 и начинается с быстрой реверсии мембранного потенциала от ПП (примерно - 90 мВ) до потенциала инверсии (пример")
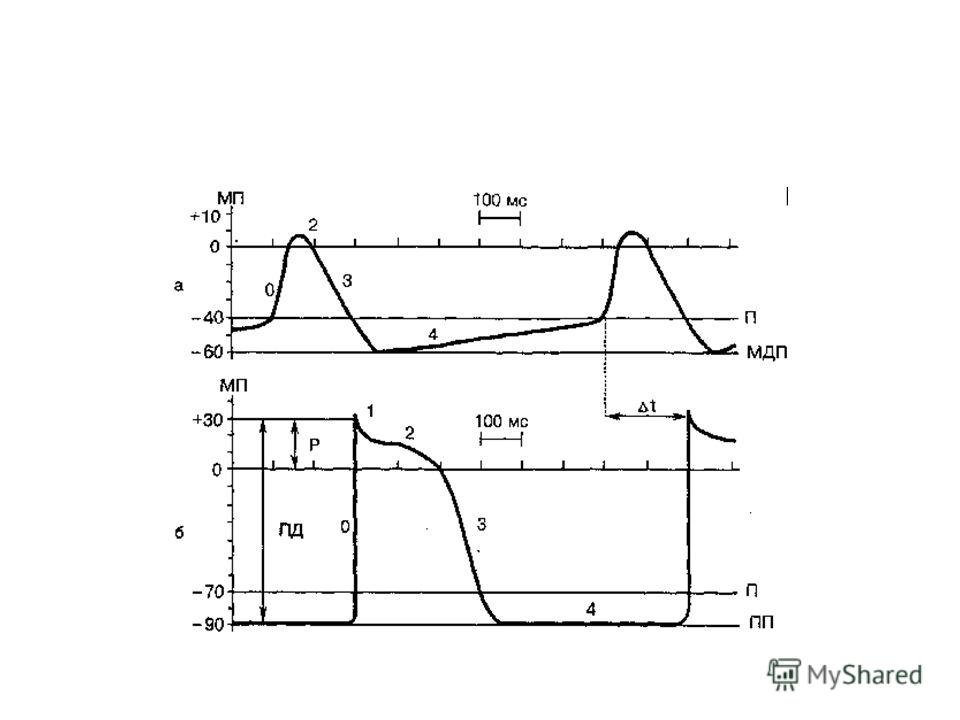
58
За этой фазой быстрой деполяризации (продолжительность – 1-2 мс) следует более длительная фаза плато – специфическая особенность клеток миокарда, затем наступает фаза реполяризации, по окончании которой восстанавливается ПП.
 следует более длительная фаза плато – специфическая особенность клеток миокарда, затем наступает фаза реполяризации, по окончании которой восстанавливается ПП.")
59
Длительность ПД кардиомиоцитов составляет мс, т.е. более чем в 100 раз превышает соответствующую величину для скелетных мышц и нервных волокон.

61
ПП близок к К + -равновесному потенциалу; деполяризация обусловлена лавинообразно нарастающим Na + -током (однако, этот Na-ток быстро инактивируется); фаза плато обусловлена входящим Са 2+ -током (медленный входящий ток) + снижение проводимости для К +, возникающее при деполяризации и уменьшающее реполяризацию; реполяризация обусловлена выходящим К + -током и снижением проводимости для иона Са 2+.
; фаза плато обусловлена входящим Са 2+ -током (медленный входящий ток) + снижение проводимости для К")
62
Специфическая форма ПД ТМВ имеет большое функциональное значение, так как определенным фазам ПД соответствует определенные изменения возбудимости мембраны (фазы рефрактерности),
,")
63
Во время длительной деполяризации мембраны (плато) Na + -каналы инактивируются, и ТМВ находится в состоянии абсолютной рефрактерности. Восстановление активности натриевых каналов происходит только после снижения МП до уровня, примерно равного – 40 мВ.
 Na + -каналы инактивируются, и ТМВ находится в состоянии абсолютной рефрактерности. Восстановление активности натриевых каналов происходит только после снижения МП до уровня, примерно равного – 40 мВ")
64
Длительный рефрактерный период предохраняет сердце от слишком быстрого повторного возбуждения и повторного сокращения. Такое возбуждение, возникшее до расслабления мышечного волокна, могло бы привести к нарушению нагнетательной функции сердца (тетанус миокарда).

65
ПД атипичных мышечных волокон – отличается отсутствием устойчивого уровня ПП. Эти клетки спонтанно деполяризуются до критического уровня. Фазы ПД – медленная диастолическая деполяризация до КМП; быстрая ДП; более или менее выраженная фаза плато; быстрая реполяризация.

67
4. Проводящая система сердца. Распространение возбуждения по миокарду. АТМВ миокарда образуют так называемую проводящую систему. Она представляет собой совокупность узлов и пучков атипичной мышечной ткани, функцией которой является генерация ПД, служащих стимулами для ТМВ, то есть задание определенного ритма сердечных сокращений.

68
Строение проводящей системы обеспечивает строго согласованное и последовательное возбуждение и сокращение различных отделов сердца.

70
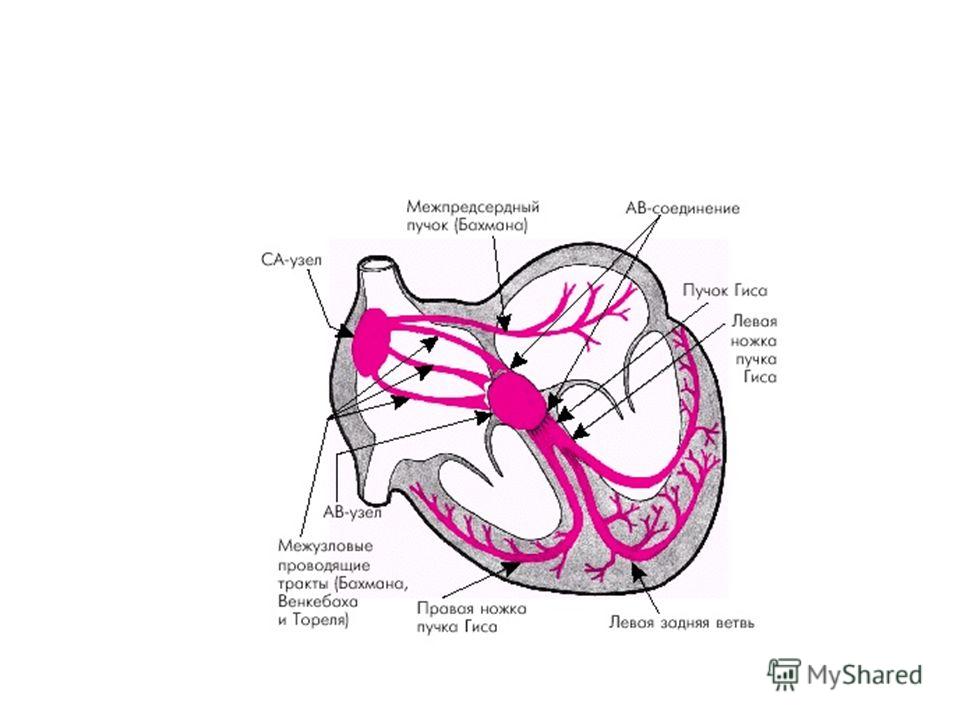
В норме водителем ритма является синоатриальный узел, расположенный в стенке правого предсердия в месте впадения в него верхней полой вены. Частота разрядов СА в покое составляет около 70 1/мин. От этого узла возбуждение вначале распространяется по рабочему миокарду предсердий (со скоростью порядка 1 м/с).

72
Единственный путь, по которому возбуждение может пройти к желудочкам, образует атриовентрикулярный узел (АВ), лежащий в предсердно-желудочковой перегородке (остальная часть атриовентрикулярного соединения образована невозбудимой соединительной тканью). В АВ узле скорость проведения значительно падает (в раз; 0,02-0,05 м/с) за счет снижения диаметра волокон АВ-узла и поперечного их расположения
, лежащий в предсердно-желудочковой перегородке (остальная часть атриовентрикулярного соединения образована невозбудимой соединительной тканью")
73
Это приводит к тому, что возбуждение "задерживается" в АВ-узле (АВ-задержка необходима для полного перехода крови из предсердий в желудочки во время сокращения предсердий).
.")
74
Далее возбуждение распространяется по пучку Гиса, ножкам пучка Гиса и волокнам Пуркинье к верхушке сердца со все возрастающей (до 4-5 м/с) скоростью (увеличение диаметра АТМВ), а затем переходит на рабочие волокна миокарда, по которым распространяется в обратном направлении – от верхушки сердца к основанию. За волной возбуждения следует сокращение ТМВ миокарда.
 скоростью (увеличение диаметра АТМВ), а затем переходит на рабочие волокна миокарда, по которым распространяетс")
75
5. Электрокардиография. Электрокардиограмма. Интегральный электрический вектор сердца. Сложный характер распространения возбуждения по сердцу отображается в электрокардиограмме (ЭКГ), по форме которой можно судить о возбудимости и проводимости различных отделов сердца (но не о сократимости волокон миокарда!)
, по форме которой можно судить о возбудимости и проводимости различ")
76
Если рассмотреть отдельное миокардиальное волокно, то в покое его наружная поверхность имеет положительный, а внутренняя – отрицательный потенциал. При возбуждении (ПД) возбужденный участок мембраны меняет свою полярность (т.е. снаружи – «-», а внутри «+»).
 возбужденный участок мембраны меняет свою полярность (т.е. снаружи – «-», а внутр")
77
Возбужденное волокно можно рассматривать как диполь, обладающий определенным дипольным моментом. Векторная сумма дипольных моментов всех волокон миокарда называется интегральным электрическим вектором сердца (ИЭВС).
.")
78
Этот вектор в каждый момент времени направлен от наиболее возбужденного (электроотрицательного) к наименее возбужденному (электроположительному) участку сердца, и величина и направление его в ходе сердечного цикла многократно меняются.
 к наименее возбужденному (электроположительному) участку сердца, и величина и направление его в ходе сердечного цикла многократно меняются.")
79
Как известно, движущиеся заряды создают вокруг себя переменное электрическое поле, которое распространяется в пространстве. Поэтому работающее сердце также является источником электрического поля, которое можно зарегистрировать на поверхности тела.

80
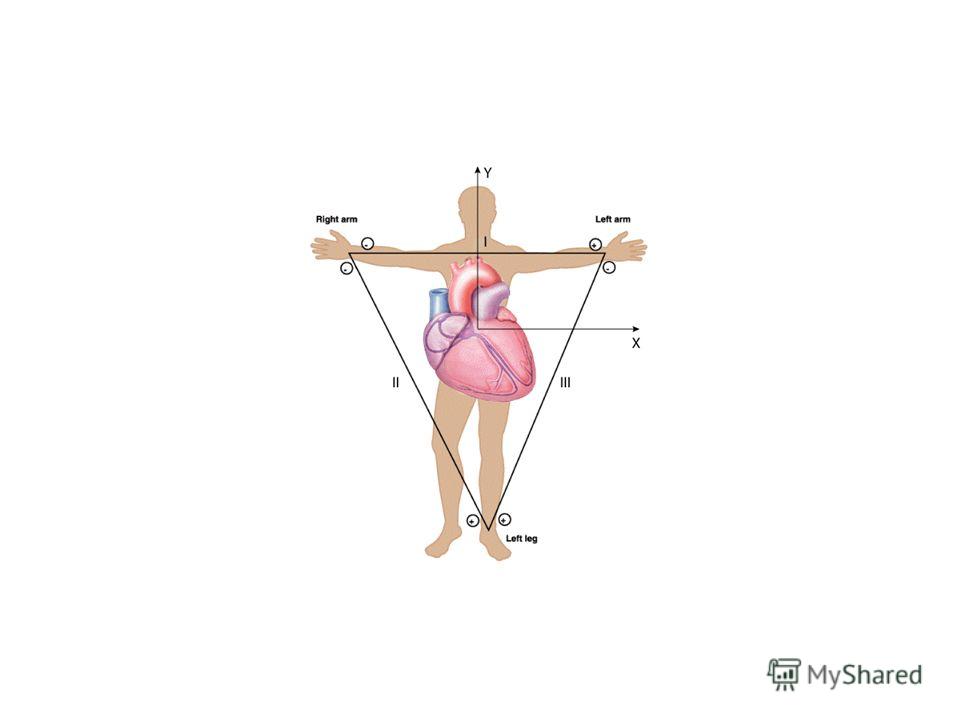
Для этого на различные точки поверхности тела накладывают отводящие электроды и регистрируют разность потенциалов между ними. Регистрирующий прибор (электрокардиограф) по сути представляет собой усилитель переменного тока и регистрирующее устройство (самописец).
 по сути представляет собой усилитель переменного тока и регистрирующее устройство")
82
Кривая, отображающая зависимость этой разности потенциалов от времени, называется электрокардиограммой. Она представляет собой периодическое (Т = 1/ЧСС) колебание сложной формы.
 колебание сложной формы.")
84
Величина разности потенциалов, регистрируемой между двумя электродами, находящимися на поверхности тела человека будет зависеть от величины интегрального электрического вектора и угла между направлением этого вектора и осью отведения (проведенной между этими электродами). Таким образом, ЭКГ представляет собой динамику во времени проекции ИЭВС на ось отведения.

85
Виллем Эйнтховен ( ), Нобелевский лауреат 1924 г.
, Нобелевский лауреат 1924 г.")
Еще похожие презентации в нашем архиве:











 Лекция 2 ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ План лекции:")










 и предполагаемая модель расположения в мембране его а1 -субъединицы.")

. 2. Мембранный потенциал (МП). 3. Потенциал действия.")



