Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
ГЛЮКОНЕОГЕНЕЗ

2
Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пировиноградная кислоты, гликогенные аминокислоты, глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот.

4
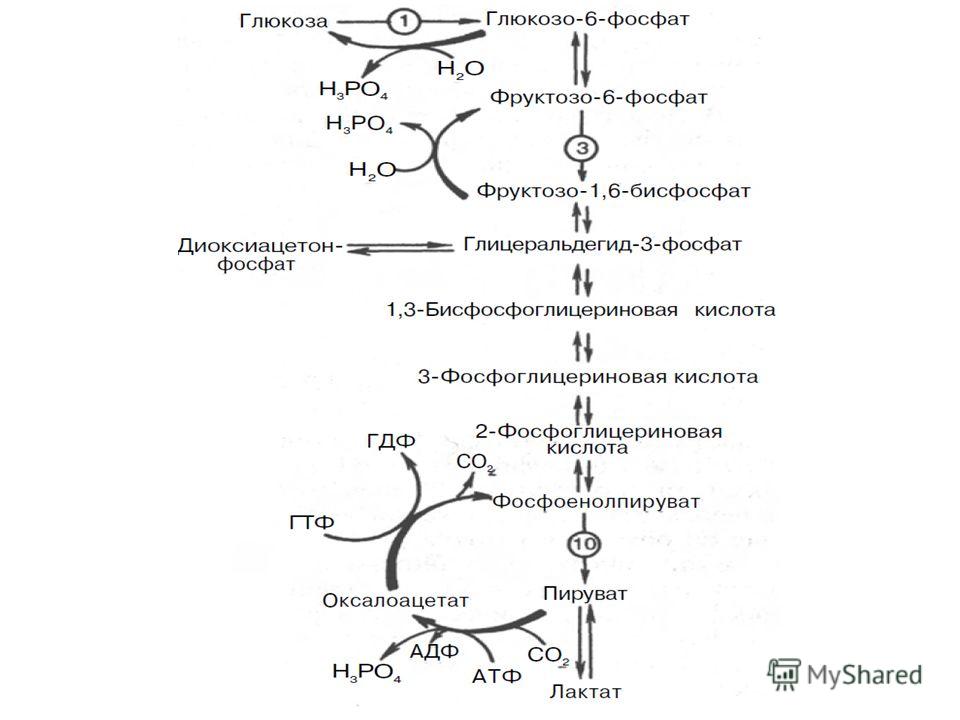
У позвоночных наиболее интенсивно глюконеогенез протекает в клетках печени и почек (в корковом веществе). Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза. Только 3 реакции гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) необратимы, поэтому в процесс глюконеогенеза на 3 этапах используются другие ферменты.
. Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза. Только 3 реакции гликолиза (гексокиназная, фосфофруктокиназная")
5
Синтез фосфоенолпирувата осуществляется в несколько этапов: 1) Превращение пирувата в оксалоацетат. Пируват карбоксилируется пируваткарбоксилазой при участии АТФ : Пируваткарбоксилаза, которая катализирует эту реакцию, является аллостерическим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-КоА.
 Превращение пирувата в оксалоацетат. Пируват карбоксилируется пируваткарбоксилазой при участии АТФ : Пируваткарбоксилаза, которая катализирует эту реакцию, является аллостерическим митохо")
6
Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же, в митохондриях, восстанавливается в малат, который легко выходит из митохондрии через митохондриальную мембрану:

7
В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-зависимой малатдегидрогеназы:

8
Затем оксалоацетат в результате декарбоксилирования и фосфорилирования под влиянием фермента фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват в цитозоле клетки. Донором фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ):
:")
9
Фосфоенолпируват, образовавшийся из пирувата, в результате ряда обратимых реакций гликолиза превращается во фруктозо- 1,6-бисфосфат. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой реакции. Превращение фруктозо-1,6-бис-фосфата во фруктозо-6-фосфат катализируется специфической фосфатазой:

10
В последующей обратимой стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат в реакции, катализируемой глюкозо-6-фосфат-изомеразы. Последний может дефосфорилироваться под влиянием фермента глюкозо-6-фосфатазы:

12
Регуляция глюконеогенеза. Роль аллостерического активатора пируваткарбоксилазы выполняет ацетил-КоА. В отсутствие ацетил-КоА фермент почти полностью лишен активности. Когда в клетке накапливается митохондриальный ацетил-КоА, биосинтез глюкозы из пирувата усиливается. Известно, что ацетил- КоА одновременно является отрицательным модулятором пируватдегидрогеназного комплекса. Накопление ацетил-КоА замедляет окислительное декарбоксилирование пирувата, что также способствует активации глюконеогенеза.

13
Другой важный момент в регуляции глюконеогенеза – реакция, катализируемая фруктозо-1,6-бисфосфатазой – ферментом, который ингибируется АМФ. Противоположное действие АМФ оказывает на фосфофрукто киназу, т. е. для этого фермента он является аллостерическим активатором. При низкой концентрации АМФ и высоком уровне АТФ происходит стимуляция глюконеогенеза. Напротив, когда величина отношения АТФ/АМФ мала, в клетке наблюдается расщепление глюкозы. Глюконеогенез и гликолиз регулируются реципрокно, так что, если активность одного из путей относительно понижается, то активность другого пути повышается.

14
Фруктозо-2,6-бисфосфат активирует фосфофруктокиназу и ингибирует фруктозо-1,6-бисфосфатазу. Повышение в клетке уровня фруктозо-2,6-бисфосфата способствует усилению гликолиза и уменьшению скорости глюконеогенеза.

15
Фруктозо-2,6-бисфосфат это метаболит, образующийся из фруктозо-6-фосфата и выполняющий только регуляторные функции. Образование фруктозо-2,6-бисфосфата путем фосфорилирования фруктозо-6-фосфата катализирует бифункциональный фермент (БИФ), который катализирует также и обратную реакцию. В реакции фосфорилирования фруктозо-6-фосфата с использованием АТФ БИФ проявляет киназную активность, а при дефосфорилировании образованного фруктозо-2,6-бисфосфата фосфатазную. Это обстоятельство и определило название фермента бифункциональный.
, который кат")
16
Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается. При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результатом является снижение количества фруктозо-2,6- бисфосфата
. Дефосфорилированная форма БИФ характерна для периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увели")
18
Глюконеогенез может регулироваться и непрямым путем. Фермент гликолиза пируваткиназа существует в 2 формах – L и М. Форма L (от англ. liver – печень) преобладает в тканях, способных к глюконеогенезу. Эта форма ингибируется избытком АТФ и некоторыми аминокислотами, в частности аланином. М-форма (от англ. muscle – мышцы) такой регуляции не подвержена. В условиях достаточного обеспечения клетки энергией происходит ингибирование L-формы пируваткиназы. Как следствие ингибирования замедляется гликолиз и создаются условия, благоприятствующие глюконеогенезу.
 преобладает в тканях, способных к глюконеогенезу. Эта форма ингибируется избытком АТФ и некоторыми а")
20
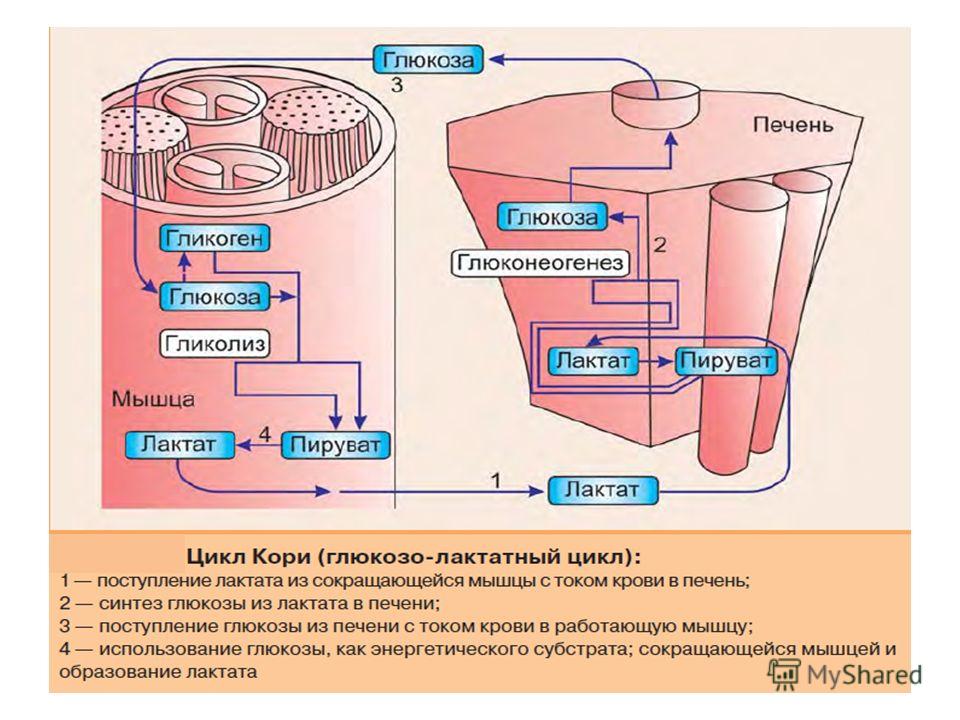
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NАDН/NАD+ ниже, чем в сокращающейся мышце, поэтому лактатдегидрогеназная реакция протекает в обратном направлении, т.е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эта последовательность событий называется глюкозо-лактатным циклом, или циклом Кори

21
Окислительное декарбоксилирование пировиноградной кислоты

22
Пируват + НАД+ + HS-KoA –> Ацетил-КоА + НАДН + Н+ + СO2 Окисление пирувата до ацетил-КоА происходит при участии мультиферментной системы, получившую название пируватдегидрогеназный комплекс Образовавшийся в процессе окислительного декарбоксилирования ацетил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбоксилирование пирувата, происходит в митохондриях клеток

23
Е1 - пируватдегидрогеназа; Е2 - дигидролипоилацетилтрансфсраза; Е3 - дигидролипоилдегидрогеназа Коферменты: ТПФ, амид липоевой кислоты, коэнзим А, ФАД, НАД стадии процесса

24
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА)
")
25
Цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль клеточного топлива : углеводов, жирных кислот и аминокислот. Цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций
, в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль клеточного топлива : углеводов, жирных кислот и аминокислот. Цикл происх")
26
Первая реакция катализируется ферментом цитрат-синтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуется лимонная кислота:

27
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис- аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат). Катализирует эти обратимые реакции гидратации–дегидратации фермент аконитатгидратаза (аконитаза).
. Катализирует эти обратимые реакции гидр")
28
Третья реакция лимитирует скорость цикла Кребса. Изолимонная кислота дегидрируется в присутствии НАД- зависимой изоцитратдегидрогеназы: НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg2+ или Мn2+

29
Во время четвертой реакции происходит окислительное декарбоксилирование α-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-КоА. Механизм этой реакции сходен с таковым реакции окислительного декарбоксилирования пирувата до ацетил-КоА. α-Кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в реакции принимают участие 5 коферментов: ТПФ, амид липоевой кислоты, HS-KoA, ФАД и НАД+:

30
Пятая реакция катализируется ферментом сукцинил-КоА- синтетазой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА: АТФ Субстратное фофорилирование
. Одновременно происходит образование высокоэргической фосфатной свя")
31
В результате шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД. В свою очередь сукцинатдегидрогеназа прочно связана с внутренней митохондриальной мембраной:
 связан кофермент ФАД. В свою очередь сукцинатдегидрогеназа прочно связа")
32
Седьмая реакция осуществляется под влиянием фермента фумаратгидратазы (фумаразы). Фумаровая кислота гидратируется, продуктом реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью: в ходе реакции образуется L-яблочная кислота:
. Фумаровая кислота гидратируется, продуктом реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью: в ходе реакц")
33
В ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:

34
Одна молекула НАДН (3 молекулы АТФ) образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление молекулы пирувата до СО2 и Н2О дает 15 молекул АТФ). К этому количеству надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 6 молекул АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН, которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции гликолиза. Следовательно, при расщеплении в тканях одной молекулы глюкозы синтезируется 38 молекул АТФ. Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем анаэробный гликолиз.
 образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цик")
35
Молекулы внемитохондриального НАДН не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицеролфосфатного челночного механизма При этом в результате полного окисления одной молекулы глюкозы может образоваться 36 молекул АТФ С помощью данного челночного механизма лишь в скелетных мышцах и мозге осуществляется перенос восстановленных эквивалентов от цитозольного НАДН + Н+ в митохондрии.

36
Глицеролфосфатный челночный механизм

37
В клетках печени, почек и сердца действует более сложная малат-аспартатная челночная система. Действие такого челночного механизма становится возможным благодаря присутствию малатдегидрогеназы и аспартатаминотрансферазы как в цитозоле, так и в митохондриях. Если функционирует малат-аспартатный механизм, то в результате полного окисления одной молекулы глюкозы может образоваться не 36, а 38 молекул АТФ

38
Малат-аспартатная челночная система

39
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ

40
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Дикенсу и В.А. Энгельгарду У млекопитающих активность пентозофосфатного цикла относительно высока в печени, надпочечниках, эмбриональной ткани и молочной железе в период лактации. Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина и т.д. За счет пентозофосфатного цикла примерно на 50% покрывается потребность организма в НАДФН Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и не участвует в окислительном фосфорилировании, протекающем в митохондриях. Пентозофосфатный цикл поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов.

41
Пентозофосфатный цикл начинается с окисления глюкозо-6- фосфата и последующего окислительного декарбоксилирования продукта (в результате от гексозофосфата отщепляется первый атом углерода). Это первая, так называемая окислительная, стадия пентозофосфатного цикла.
. Это первая, так называемая окислительная, стадия пентозофосф")
43
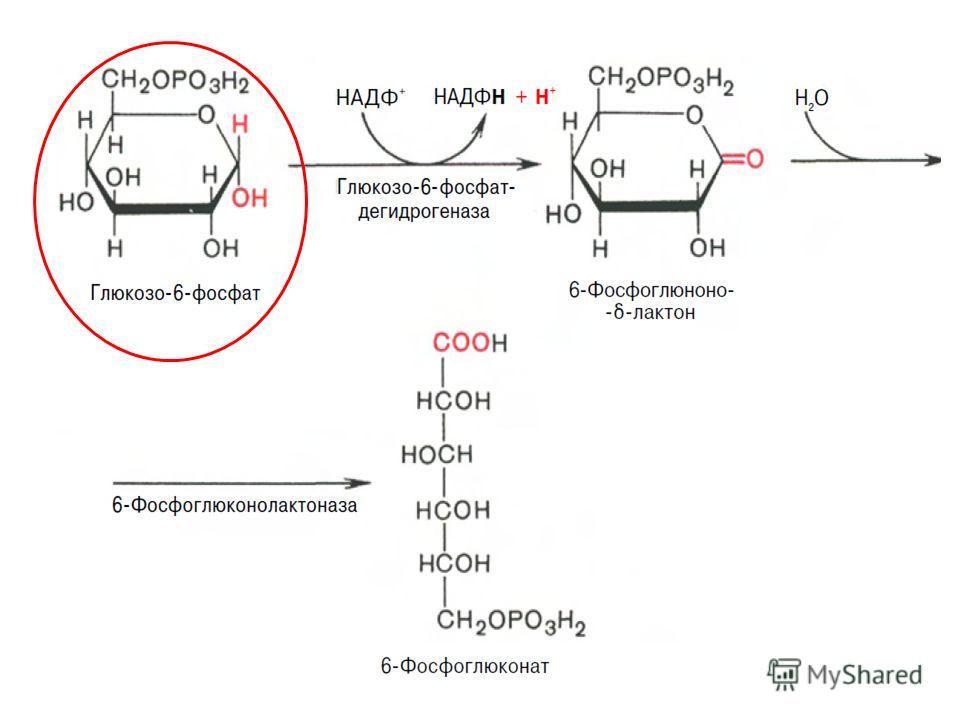
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ+. Образовавшийся в ходе реакции 6-фосфоглюконо-δ-лактон – соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фосфоглюконовой кислоты (6-фосфоглюконат) и НАДФН :

44
Во второй – окислительной – реакции, катализируемой 6-фосфоглюконатдегидрогеназой (декарбоксилирующей), 6- фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН:
, 6- фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула")
45
Под действием соответствующей эпимеразы из рибулозо-5- фосфата может образоваться другая фосфопентоза – ксилулозо-5- фосфат. Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:

46
Неокислительный этап (стадия) пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо- 6-фосфат, фруктозо-1,6-бисфосфат, фосфотриозы), а другие – специфические для пентозофосфатного пути (седогептулозо-7- фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
 пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо- 6-фосфат, фруктозо-1,")
47
Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов. Коферментом в транскетолазной реакции служит ТПФ, играющий роль промежуточного переносчика гликольальдегидной группы от ксилулозо-5-фосфата к рибозо-5-фосфату. В результате образуется семиуглеродный моносахарид седогептулозо-7-фосфат и глицеральдегид-3-фосфат:

48
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз – при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритрозо-4-фосфатом:

49
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седогептулозо-7-фосфата на глицеральдегид-3-фосфат:
 от седогептулозо-7-фосфата на глицеральдегид-3-фосфат:")
50
Пентозофосфатный путь и гликолиз, протекающие в цитозоле, взаимосвязаны и способны переключаться друг на друга в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке

51
Синдром Вернике-Косакова (нервно-псих. заболевание) связан со значительным снижением (в 10 раз) способности транскетолазы связывать кофермент ТПФ. Дефект гена глюкозо-6- фосфатдегидрогеназы в эритроцитах сопровождается гемолитической анемией. Причина – недостаток НАДФН и, как следствие, недостаток восстановленного глутатиона (GSH), что приводит к росту образования активных форм кислорода и гемолизу эритроцитов
 связан со значительным снижением (в 10 раз) способности транскетолазы связывать кофермент ТПФ. Дефект гена глюкозо-6- фосфатдегидрогеназы в эритроцитах сопровождается гемолитической анемией. Причина")
52
Типы гликогенозов и их характеристика

Еще похожие презентации в нашем архиве:




 (Mg 2+ ) (2.7.1.1.) 2.Глюкозофосфатизомераза (5.3.1.9.)")

 и синтез глюкозы (глюконеогенез) Автор – доцент кафедры биохимии Рыскина Е.А.")

 Пластический обмен (анаболизм, ассимиляция) Совокупность реакций синтеза, которые идут с затратой энергии АТФ.")



подготовительный.")


 Совокупность протекающих в клетке химических превращений, обеспечивающих её рост,")


 Кислородный ( аэробный)")





