Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
СИНТЕЗ ВЫСШИХ ЖИРНЫХ КИСЛОТ, ФОСФОЛИПИДОВ, СФИНГОМИЕЛИНА, ХОЛЕСТЕРИНА. ПРЕВРАЩЕНИЕ ГЛИЦЕРИНА

2
Синтез высших жирных кислот Основную регуляторную реакцию синтеза ВЖК катализирует биотин- содержащий фермент ацетил-КоА-карбоксилаза, в ходе которой ацетил- КоА превращается в малонил-КоА. Реакция протекает в два этапа: I – карбоксилирование биотина с учас тием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в ре зультате чего образуется малонил-КоА:

3
Благодаря ключевому положению этой реакции в синтезе ВЖК активность ацетил-КоА-карбоксилазы может изменяться в широких пределах путем: – ассоциации и диссоциации протомеров. Цитрат стимулирует ассоциацию и повышает активность фермента, а увеличение концентрации ацил-КоА ускоряет диссоциацию протомеров и снижает активность фермента. – фосфорилирования и дефосфорилирования. Инсулин стимулирует дефосфорилирование и повышает активность фермента, а глюкагон и адреналин фосфорилирование и его инактивацию. – индукции синтеза новых молекул фермента под влиянием инсулина.

4
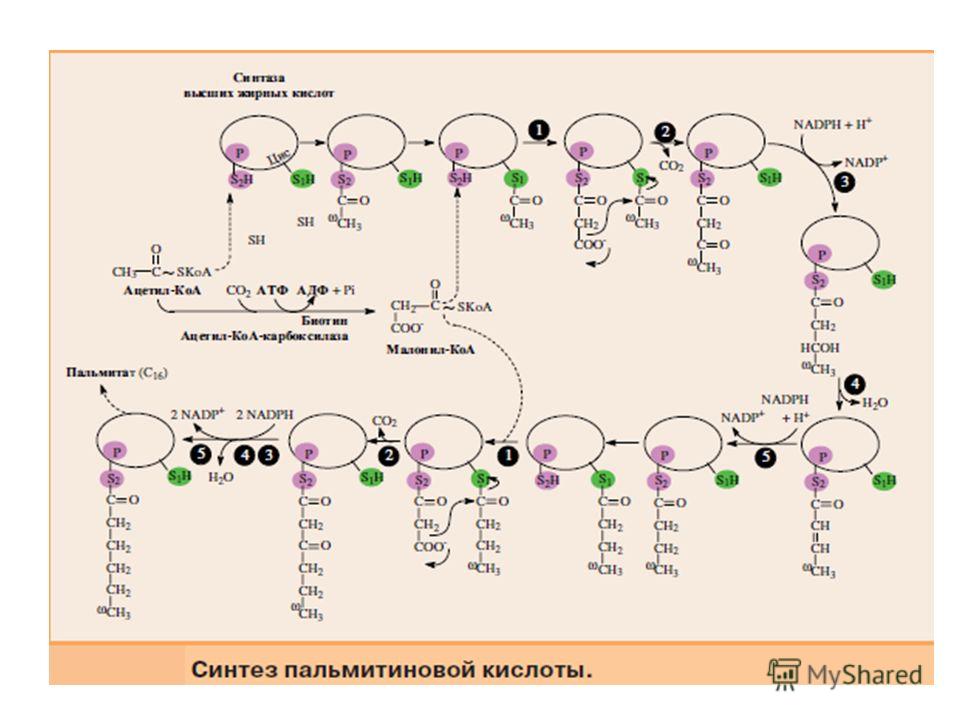
Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах. Мол. масса АПБ составляет около Данный белок в синтетазной системе выполняет роль КоА. Последовательность реакций, происходящих при синтезе жирных кислот:
 жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы и вовлекается в процесс с")
6
Далее цикл реакций повторяется. Допустим, что идет синтез пальмитиновой кислоты (С16). В этом случае образованием бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы малонил- АПБ к карбоксильному концу растущей цепи жирной кислоты. При этом отщепляется дистальная карбоксильная группа малонил-АПБ в виде СО2. Например, образовавшийся в первом цикле бутирил-АПБ взаимодействует с малонил-АПБ:
. В этом случае образованием бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы малонил- АПБ к карбоксильно")
7
Завершается синтез жирной кислоты отщеплением HS-АПБ от ацил- АПБ под влиянием фермента деацилазы.

8
Синтез пальмитиновой кислоты

10
По сравнению с β-окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитозоле клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путем связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ); при биосинтезе образуется D(–)-изомер 3-гидроксикислоты, а не L(+)- изомер, как это имеет место при β-окислении жирных кислот; необходимость для синтеза жирных кислот кофермента НАДФН. НАДФН в организме частично (на 50%) образуется в реакциях пентозофосфатного цикла, частично – в других реакциях, в частности в реакциях: Малат + НАДФ+ –> Пируват + СO2 + НАДФН + Н+ Изоцитрат + НАДФ+ –> α-Кетоглутарат + СO2 + НАДФН + Н+.

11
Жирные кислоты с числом углеродных атомов больше, чем 16, (элонгация) синтезируются в эндоплазматическом ретикулуме аналогично тому, как это происходит на пальмитилсинтазе. Однако каждую стадию процесса катализируют отдельные ферменты. Удлинение цепи происходит с помощью малонил-КоА, а в реакциях восстановления используется NADPH + H+. Образование ненасыщенных жирных кислот (десатурация) пальмитоолеиновой и олеиновой кислот происходит на мембране эндоплазматического ретикулума (ЭР), двойные связи между С9 и С10 возникают за счет работы оксигеназы жирных кислот, которая требует для своей работы участия О2 и NADPH. Синтез жирных кислот с большем чем С16, числом углеродных атомов (элонгация)
 синтезируются в эндоплазматическом ретикулуме аналогично тому, как это происходит на пальмитилсинтазе. Однако каждую стадию процесса катализируют отдельные ферменты. Удлинение цепи")
12
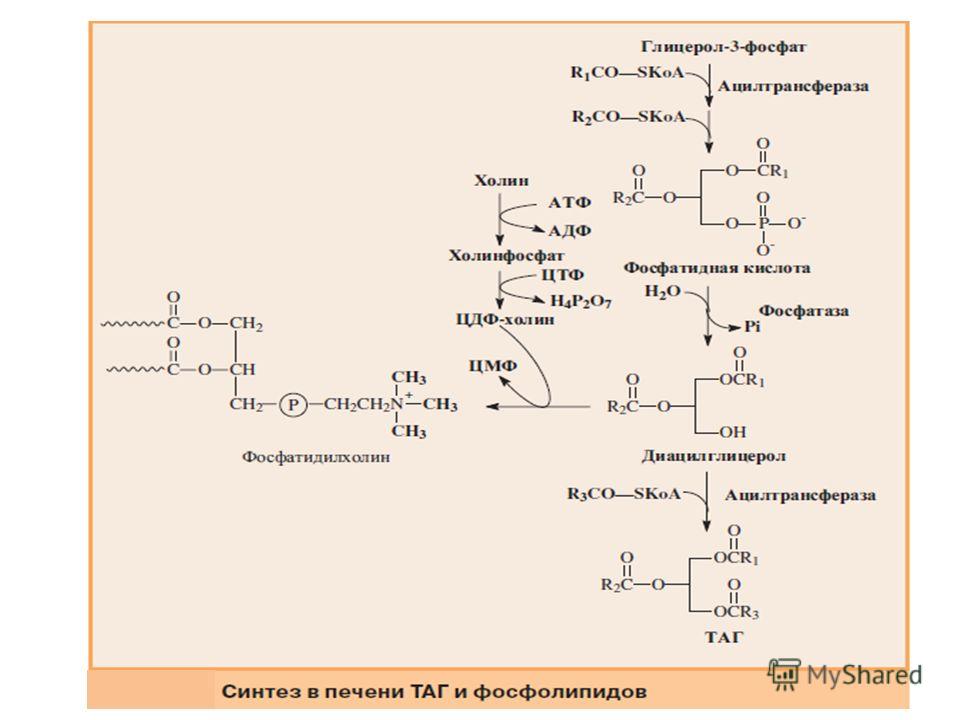
Биосинтез фосфолипидов Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сети клетки. Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов), сфингозин (в синтезе сфингомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов.

13
Биосинтез фосфатидилэтаноламина Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфоэтаноламина:

14
Затем фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (PPi):
 и пирофосфат (PPi):")
15
В следующей реакции ЦДФ-этаноламин, взаимодействуя с 1,2-диглицеридом, образующимся при дефосфорилировании фосфатидной кислоты, превращается в фосфатидилэтаноламин. Реакция катализируется ферментом этаноламинфосфотрансферазой:

16
Биосинтез фосфатидилхолина (лецитина) Фосфатидилэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метальных групп от трех молекул S-аденозилметионина (донор метальных групп) к аминогруппе остатка этаноламина образуется фосфатидилхолин:
 Фосфатидилэтаноламин является предшественником фосфатидилхолина. В результате последовательного переноса трех метальных групп от трех молекул S-аденозилметионина (донор метальных групп) к аминогруппе остатка этан")
17
Существует еще один путь синтеза фосфатидилхолина в клетках животных. В этом случае, как и при синтезе фосфатидилэтаноламина, используется ЦТФ в качестве переносчика, но уже не фосфоэтаноламина, а фосфохолина. На первом этапе синтеза свободный холин активируется под действием холинкиназы с образованием фосфохолина:

19
Биосинтез фосфатидилсерина У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин:

20
Существует и второй путь образования фосфатидилсерина, который связан с предварительным вовлечением фосфатидной кислоты в синтез фосфоглицеридов. Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина.

21
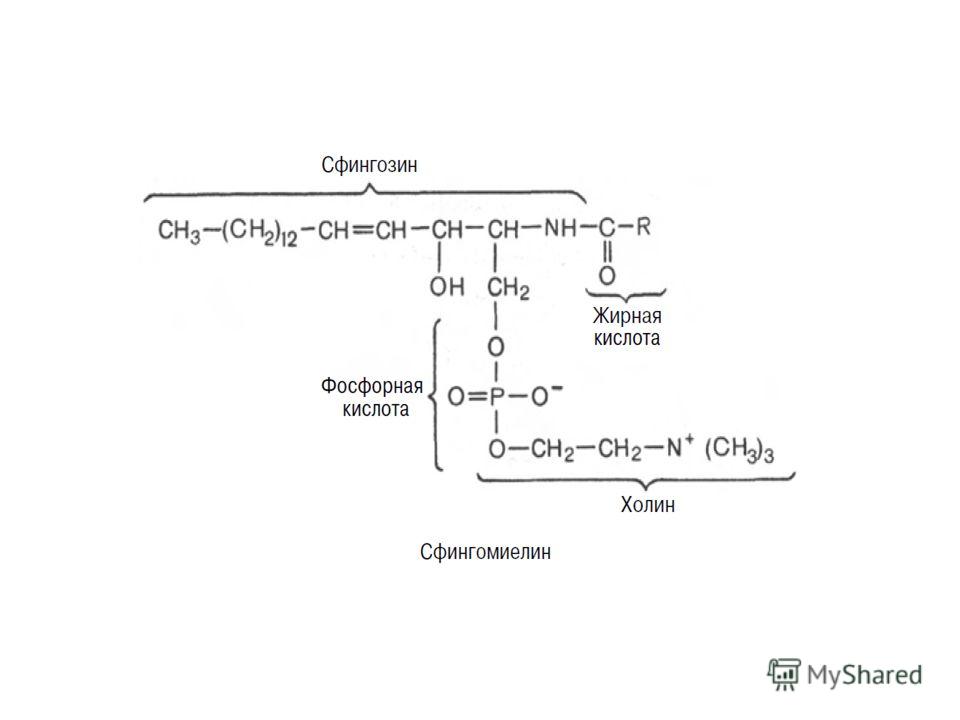
Биосинтез сфингомиелина Интермедиатом в биосинтезе сфингомиелина является церамид (N-ацилсфингозин), который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:
, который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:")
23
БИОСИНТЕЗ ХОЛЕСТЕРИНА В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в мевалоновую кислоту, II – образование сквалена из мевалоновой кислоты, III – циклизация сквалена в холестерин I стадия Начальным этапом синтеза мевалоновой кислоты из ацетил-КоА является образование ацетоацетил-КоА посредством обратимой тиолазной реакции:

24
В результате конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при участии гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА-синтаза) образуется β-гидрокси-β-метилглутарил-КоА:
 образуется β-гидрокси-β-метилглутарил-КоА:")
25
β-Гидрокси-β-метилглутарил-КоА под действием регуляторного фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА- редуктаза) в результате восстановления одной из карбоксильных групп и отщепления HS-KoA превращается в мевалоновую кислоту: ГМГ-КоА-редуктазная реакция – первая практически необратимая реакция в цепи биосинтеза холестерина. Активность ГМГ-редуктазы возрастает при введении инсулина и тиреоидных гормонов. Это приводит к усилению синтеза холестерина и повышению его уровня в крови. При голодании, тиреоидэктомии, введение глюкагона и глюкокортикоидов, напротив, отмечается угнетение синтеза холестерина, что прежде всего связано со снижением активности ГМГ-КоА-редуктазы.
 в результате восстановления одной из карбоксильных групп и отщепления HS-KoA превращается в мевалоновую кислот")
26
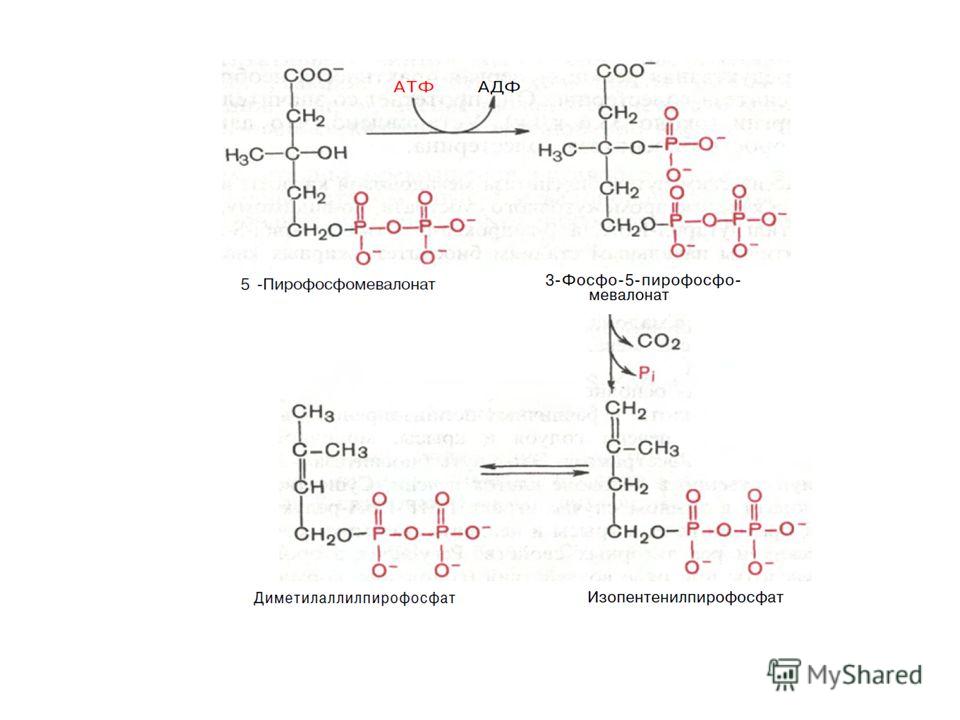
II стадия Мевалоновая кислота превращается в сквален. Реакции II стадии начинаются с фосфорилирования мевалоновой кислоты с помощью АТФ. В результате образуется 5-фосфорный эфир, затем 5-пирофосфорный эфир мевалоновой кислоты. Мевалонат за счет декарбоксилирования с потреблением АТФ превращается в изопентенилдифосфат, который и является тем структурным элементом, из которого строятся все изопреноидыдекарбоксилированияАТФизопреноиды

28
Оба изомерных изопентенилпирофосфата (диметилаллилпирофосфат и изопентенилпирофосфат) конденсируются с высвобождением пи- рофосфата и образованием геранилпирофосфата:
 конденсируются с высвобождением пи- рофосфата и образованием геранилпирофосфата:")
29
К геранилпирофосфату вновь присоединяется изопентенилпирофосфат. В результате этой реакции образуется фарнезилпирофосфат: Фарнезилдифосфат является также исходным соединением для синтеза других полиизопреноидов, например, убихинонаубихинон

30
В заключительной реакции данной стадии в результате НАДФН-зависимой восстановительной конденсации 2 молекул фарнезилпирофосфата образуется сквален:

31
III стадия Сквален, линейный изопреноид, под влиянием сквален-оксидоциклазы циклизируется с потреблением кислорода в ланостерин, С 30 -стерин, от которого на последующих стадиях, катализируемых цитохромом Р450, отщепляются три метильные группы, далее насыщением двойной связи в боковой цепи и перемещением двойной связи в кольце В из положения 8, 9 в положение 5, 6 вследствие чего образуется конечный продукт холестерин.изопреноидкислородацитохромом холестерин

32
ОБЩАЯ СХЕМА СИНТЕЗА ХОЛЕСТЕРИНА

33
ПРЕВРАЩЕНИЕ ГЛИЦЕРИНА 22 АТФ

34
Распад липидов в тканях осуществляют гидролитические ферменты– липазы. Три-, ди- и моноглицеридлипазы действуют на нейтральные жиры. Фосфолипазы А1, А2, С и Д (лизосомные) расщепляют кислые фосфолипиды. В результате липолиза образуются энергетические субстраты: жирные кислоты и глицерин. Глицерин в реакциях окисления (дегидрирования) и фосфорилирования (фосфокиназные) превращается в глицероальдегидфосфат. Далее путем глюнеогенеза из него синтезируются запасные энергетические субстраты (глюкоза, гликоген) или при гликолитическом окислении образуется энергия в виде молекул АТФ. Глицерол-3-фосфат может использоваться для синтеза собственных липидов.
 расщепляют кислые фосфолипиды. В результате липолиза образуются энергетические")
Еще похожие презентации в нашем архиве:



 и синтез глюкозы (глюконеогенез) Автор – доцент кафедры биохимии Рыскина Е.А.")

 Профессор, д.м.н. – Б.С. Хышиктуев.")





 Совокупность протекающих в клетке химических превращений, обеспечивающих её рост,")
 Пластический обмен (анаболизм, ассимиляция) Совокупность реакций синтеза, которые идут с затратой энергии АТФ.")




 Кислородный ( аэробный)")


 - это высокоспецифичные белки, выполняющие функции биологических катализаторов. Катализатор - это вещество, которое ускоряет.")
