Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
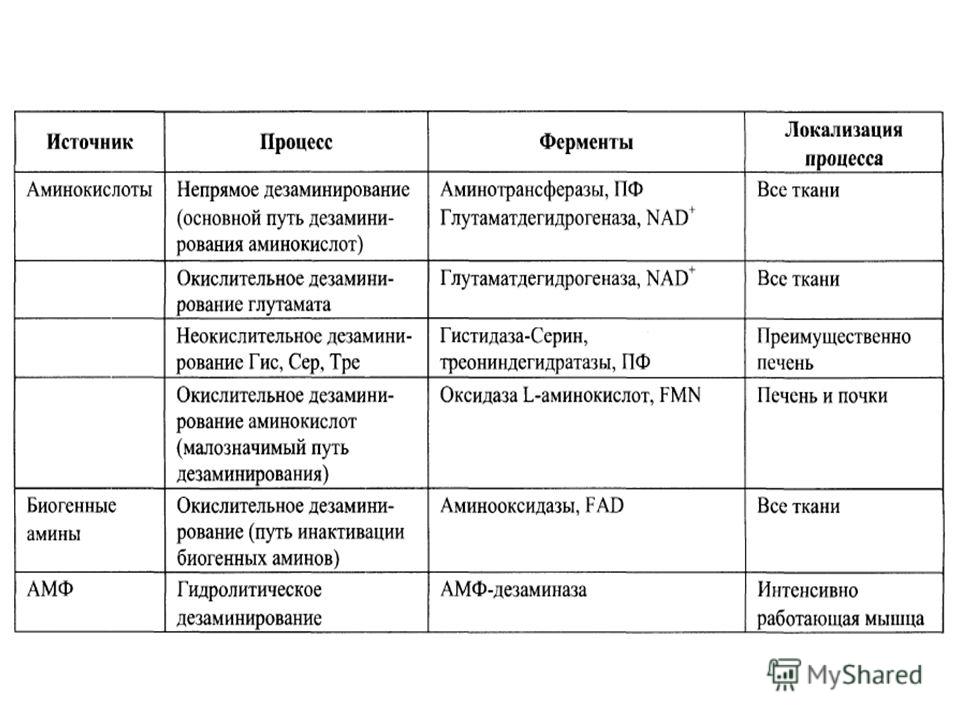
Пути образования и обезвреживания аммиака в организме

4
В организме человека подвергается распаду около 70 г АМК/сутки, при этом в результате реакций дезаминиро- вания и окисления биогенных аминов освобождается большое количество аммиака, являющегося высокотоксичным соединением. Уровень аммиака в крови в норме не превышает 60 мкмоль/л (это почти в 100 раз меньше концентрации глюкозы в крови). Концентрация аммиака 3 ммоль/л является летальной. Аммиак должен подвергаться связыванию в тканях с образованием нетоксичных соединений, легко выделяющихся с мочой

5
Пути обезвреживания аммиака

6
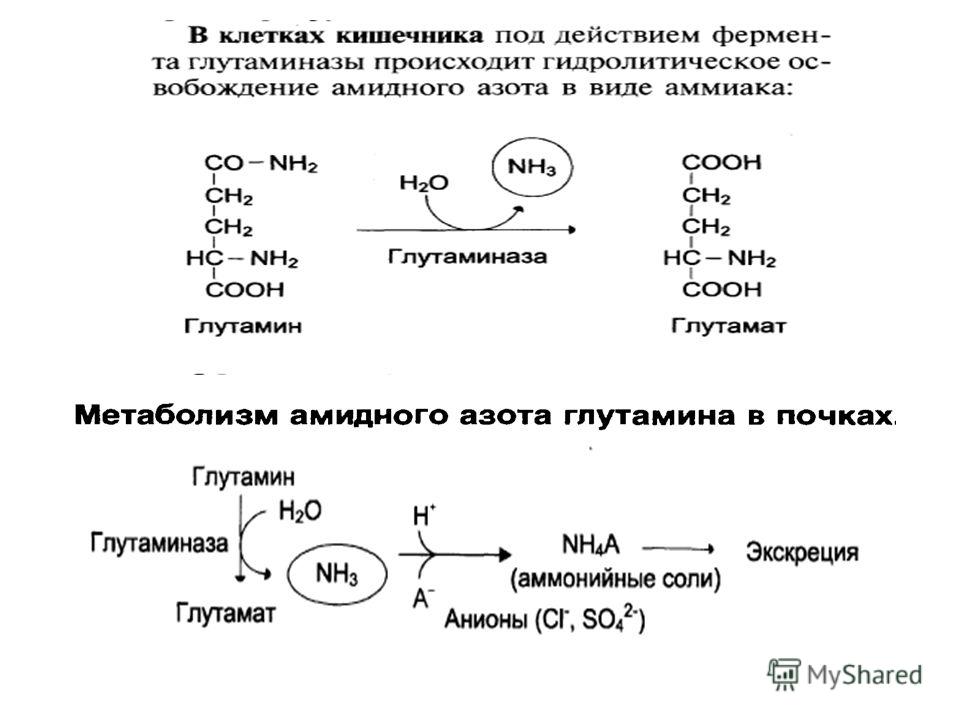
Один из путей связывания и обезвреживания аммиака в организме, в частности в мозге, сетчатке, почках, печени и мышцах,– это биосинтез глутамина (аспарагина). Глутамин и аспарагин выделяются с мочой в небольшом количестве
. Глутамин и аспарагин выделяются с мочой в небольшом количестве")
7
Cинтез глутамина, катализируемый глутаминсинтетазой: Этапы синтеза:

8
У микроорганизмов и в животных тканях открыта специфическая аммиакзависимая аспарагинсинтетаза, которая катализирует синтез аспарагина в две стадии: В животных тканях содержится, кроме того, глутаминзависимая аспарагинсинтетаза, которая для синтеза во второй стадии использует амидную группу глутамина: Суммарная ферментативная реакция синтеза аспарагина:

9
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80–85% от всего азота мочи. Основным местом синтеза мочевины является печень. Впервые Г. Кребс и К. Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего название орнитинового цикла мочевинообразования Кребса. Следует указать, что в биохимии это была первая циклическая система

10
На первом этапе синтезируется макроэргическое соединение карбамоилфосфат – метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза пиримидиновых нуклеотидов (соответственно ДНК и РНК) и аргинина (соответственно белка и мочевины):
 и аргинина (соответственно бе")
11
Открыты три разных пути синтеза карбамоилфосфата de novo, катализируемые тремя разными ферментами. Первую необратимую реакцию катализирует регуляторный фермент – аммиакзависимая карбамоилфосфатсинтетаза: Реакция требует затраты двух молекул АТФ, открыта в митохондриях клеток печени и используется преимущественно для синтеза аргинина и мочевины. В этой реакции в качестве активного стимулирующего аллостерического эффектора действует N-ацетилглутамат

12
Вторую, также необратимую, реакцию катализирует глутаминзависимая карбамоилфосфатсинтетаза: Данная реакция открыта в цитозоле клеток животных и требует наличия ионов Mg2+. Следует указать, что благодаря включению гидролитической стадии она используется преимущественно для синтеза пиримидиновых нуклеотидов. Третью обратимую реакцию катализирует карбаматкиназа: Реакция открыта у разных микроорганизмов и используется для ресинтеза АТФ.

13
Орнитиновый цикл синтеза мочевины в печени

14
Окислительное дезаминирование глутамата, синтез карбамоилфосфата и цитрулина происходят в митохондриях, а все остальные реакции цикла в цитозоле. Перенос цитруллина из митохондрий в цитозоль, и орнитина, образованного из аргинина, в митохондрии осуществляется с помощью специфической транслоказы

15
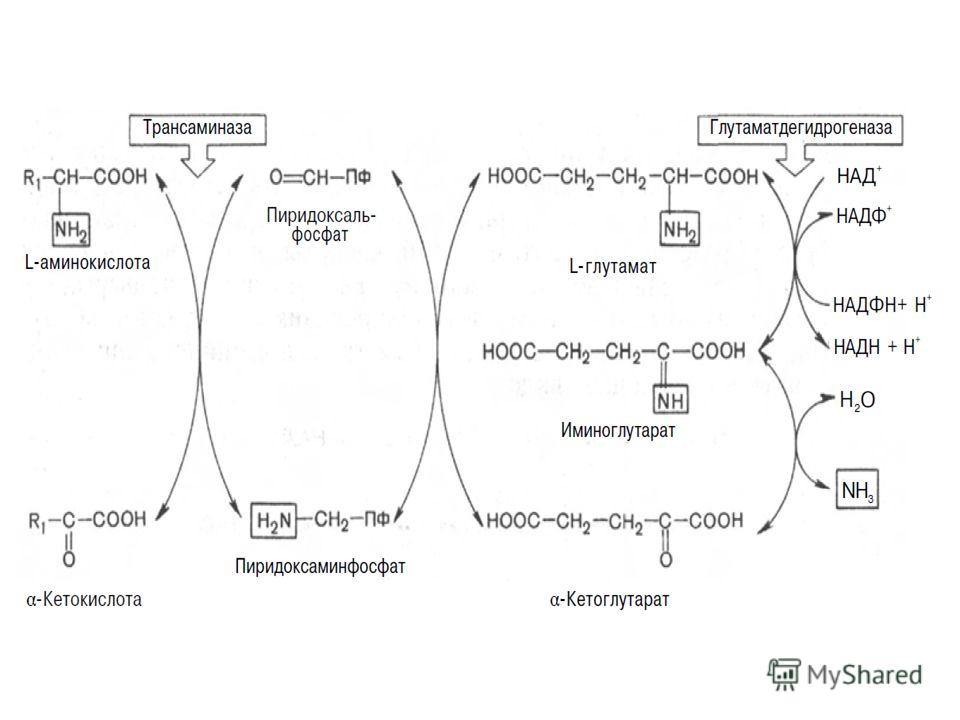
Учитывая известные фактические данные о механизмах обезвреживания аммиака в организме, можно сделать следующее заключение. Часть аммиака используется на биосинтез аминокислот путем восстановительного аминирования α-кетокислот по механизму реакции трансаминирования. Аммиак связывается при биосинтезе глутамина и аспарагина. Некоторое количество аммиака выводится с мочой в виде аммонийных солей. В форме креатинина, который образуется из креатина и креатинфосфата, выделяется из организма значительная часть азота аминокислот. Наибольшее количество аммиака расходуется на синтез мочевины, которая выводится с мочой в качестве главного конечного продукта белкового обмена в организме человека и животных.

16
В процессе эволюции живые организмы выработали различные типы азотистого обмена. Аммониотелический тип, при котором главным конечным продуктом азотистого обмена является аммиак, свойствен преимущественно рыбам. При уреотелическом типе обмена основным конечным продуктом обмена белков является мочевина; такой тип характерен для человека и животных. Урикотелический тип характерен для птиц и рептилий; главным конечным продуктом данного типа обмена является мочевая кислота.

17
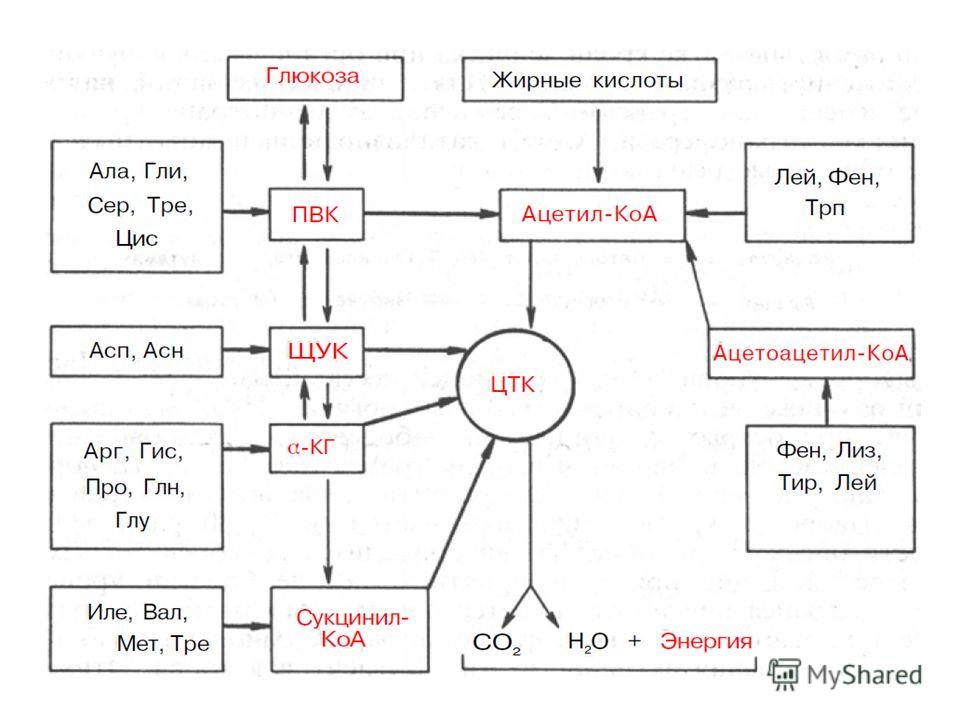
Катаболизм безазотистых остатков, полученных в результате дезаминирования аминокислот, приводит к образованию либо ацетил-КоА или к образованию метаболитов, способных включаться в глюконеогенез. Все кетокислоты способны окисляться в цитратном цикле до СО2 и Н2О с выделением энергии. Безазотистые остатки большинства аминокислот при катаболизме проходят стадию образования оксалоацетата или пирувата, превращаясь в них непосредственно либо опосредованно через промежуточные продукты цитратного цикла. Такие аминокислоты называются гликогенными, так как пируват и оксалоацетат являются субстратами глюконеогенеза.

19
Углеродные скелеты аминокислот могут включаться в ЦТК через ацетил-КоА, пируват, оксалоацетат, α-кетоглутарат и сукцинил- КоА. Пять аминокислот (Фен, Лиз, Лей, Трп, Тир) считаются кетогенными, поскольку они являются предшественниками кетоновых тел, в частности ацетоуксусной кислоты, в то время как большинство других аминокислот, обозначаемых как гликогенные, служат в организме источником углеводов, в частности глюкозы. Подобный синтез углеводов de novo усиливается при некоторых патологических состояниях, например при сахарном диабете, а также при гиперфункции коркового вещества надпочечников и введении глюкокортикоидов. Разделение аминокислот на кетогенные и гликогенные носит, однако, условный характер, поскольку отдельные участки углеродных атомов Лиз, Трп, Фен и Тир могут включаться и в молекулы предшественников глюкозы, например Фен и Тир – в фумарат. Истинно кетогенной аминокислотой является только лейцин.
 считаются кетогенными, поскольку они являются предшественниками кетоновых тел, в")
20
Механизм действия пиридоксальфосфата в аспартатаминотрансферазе

21
В связи с тем что во всех пиридоксалевых ферментах (включая трансаминазы) карбонильная группа кофермента (СНО) оказалась связанной с ε- аминогруппой лизина белковой части, в классический механизм реакции трансаминирования А.Е. Браунштейн и Э. Снелл внесли следующее дополнение. Оказалось, что взаимодействие между субстратом, т.е. L-амино-кислотой (на рисунке – аспартат), и пиридоксальфосфатом происходит не путем конденсации с выделением молекулы воды, а путем реакции замещения, при которой NH2-группа субстрата вытесняет ε-NН2-группу лизина в молекуле ферментного белка, что приводит к формированию пиридоксальфосфатного комплекса.
 карбонильная группа кофермента (СНО) оказалась связанной с ε- аминогруппой лизина белковой части, в классический механизм реакции трансаминирования А.Е. Браунштейн и Э. Снелл в")
22
Образование ПФ-субстратного комплекса, представленного шиффовым основанием ПФ и аминокислоты, в сочетании с некоторым оттягиванием электронов белковой частью молекулы фермента сопровождается лабилизацией одной из трех связей при α-углеродном атоме, благодаря чему аминокислота способна вступать в реакции трансаминирования (а), декарбоксилирования (b) и альдольного расщепления (с).

Еще похожие презентации в нашем архиве:


 и синтез глюкозы (глюконеогенез) Автор – доцент кафедры биохимии Рыскина Е.А.")




 Совокупность протекающих в клетке химических превращений, обеспечивающих её рост,")


подготовительный.")
 по теме: обмен веществ и энергии в клетке.")

 Кислородный ( аэробный)")



 – это биологически активные вещества химической.")


