Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Биоинформатика

2
Распознавание генов

3
Таблица генетического кода

4
Поиск генов если известен белок: просто

5
… или родственный белок: тоже просто

6
Генетический код: стоп-кодоны

7
Открытые рамки считывания Ген должен располагаться внутри области от стоп- кодона до следующего стоп-кодона (в той же фазе)
")
8
Сильное перепред- сказание (Aeropyrum pernix)
")
9
Генетический код: синонимы

10
Codon usage (статистика употребления кодонов) частоты кодонов отличаются от частот триплетов в некодирующих областях –различия в частотах аминокислот в белках –различия в частотах синонимичных кодонов частоты синонимичных кодонов –специфичны для генома –коррелируют с концентрациями тРНК
 частоты кодонов отличаются от частот триплетов в некодирующих областях –различия в частотах аминокислот в белках –различия в частотах синонимичных кодонов частоты синонимичных кодонов –специфичны для гено")
11
GenMark, окно 96 нт

12
Генетический код: старт-кодоны

13
Начала генов Bacillus subtilis dnaN ACATTATCCGTTAGGAGGATAAAAATG gyrA GTGATACTTCAGGGAGGTTTTTTAATG serS TCAATAAAAAAAGGAGTGTTTCGCATG bofA CAAGCGAAGGAGATGAGAAGATTCATG csfB GCTAACTGTACGGAGGTGGAGAAGATG xpaC ATAGACACAGGAGTCGATTATCTCATG metS ACATTCTGATTAGGAGGTTTCAAGATG gcaD AAAAGGGATATTGGAGGCCAATAAATG spoVC TATGTGACTAAGGGAGGATTCGCCATG ftsH GCTTACTGTGGGAGGAGGTAAGGAATG pabB AAAGAAAATAGAGGAATGATACAAATG rplJ CAAGAATCTACAGGAGGTGTAACCATG tufA AAAGCTCTTAAGGAGGATTTTAGAATG rpsJ TGTAGGCGAAAAGGAGGGAAAATAATG rpoA CGTTTTGAAGGAGGGTTTTAAGTAATG rplM AGATCATTTAGGAGGGGAAATTCAATG

14
Участок связывания рибосом dnaN ACATTATCCGTTAGGAGGATAAAAATG gyrA GTGATACTTCAGGGAGGTTTTTTAATG serS TCAATAAAAAAAGGAGTGTTTCGCATG bofA CAAGCGAAGGAGATGAGAAGATTCATG csfB GCTAACTGTACGGAGGTGGAGAAGATG xpaC ATAGACACAGGAGTCGATTATCTCATG metS ACATTCTGATTAGGAGGTTTCAAGATG gcaD AAAAGGGATATTGGAGGCCAATAAATG spoVC TATGTGACTAAGGGAGGATTCGCCATG ftsH GCTTACTGTGGGAGGAGGTAAGGAATG pabB AAAGAAAATAGAGGAATGATACAAATG rplJ CAAGAATCTACAGGAGGTGTAACCATG tufA AAAGCTCTTAAGGAGGATTTTAGAATG rpsJ TGTAGGCGAAAAGGAGGGAAAATAATG rpoA CGTTTTGAAGGAGGGTTTTAAGTAATG rplM AGATCATTTAGGAGGGGAAATTCAATG

15
Сравнительный анализ (один и тот же ген в нескольких геномах) Sty TCGCTCG--CAGCGGAAAGAGGATTACGCCCTTCGCCTGGAGGCTGTGCAGGGGC---GCCGGAGATGGGATGCATAATT Stm TCGCTCG--CAGCGGAAAGAGGATTACGCCCTTCGCCTGGAGGCTGTGCAGGGGC---GCCGGAGATGGGATGCATAATT Sen TCGCTCG--CAGCGGAAAGAGGATTACGCCCTTCGCCTGGAGGCTGTGCAGGGGC---GCCGGAGATGGGATGCATAATT Eco TTGCCCG--TGCCAGACGGCAGATTATCTCCCTGACCTGGTGGTTGCCCAGGAGGAGGGCCGGAAATAGGTTGTATCATT Kpn ----CGG--TGGCGCAGTGCCTGATGGG-CCTCGCCCTGGAGGACGGTCTGGCAT---ATCAGCAAGGGGGTGCGTCATG Ype TTGTTAGAACAGGGGAAAACGGTAAACAGTGTGGCATTAGATGTCGGTTATAGCT-----CCGCCTCTGCTTTTATCGCC * * * * * * * * * * * Sty AATTATCCTTTAAC CATAAATCTGAGCAATA-TATGCTTGGCGGCCAGATTATGGC--ACACTTGTCCGG Stm AATTATCCTTTAAC CATAAATCTGAGCAATA-TATGCCTGGCGGCCAGATTATGGC--ACACTTGTCCGG Sen AATTATCCTTTAAC CATAAATCTGAGCAATA-TATGCCTGGCGGCCAGATTATGGC--ACACTTGTCCGG Eco ACGTATCCTTATAC CTGAAATCTTCGCAAG--TATGCCTGGCCGCGAGATTATGGC--ACACTTGTCCGG Kpn ATTCATCCTTTCGATATCGCGGTGCTGGAACCAGGTGATGAGTATGCCTGGCGGCCAGATTATGGC--ACACTTCCCCAG Ype ATGTTTCAGCAAATAT CGGGTACCA-CGCCTGAGCGTTTCCGGCGGGGCAATAGTGGCTTATACTAAGCCCC * ** * * * * *** * ** **** * *** ** Sty TTAACTCTCGTT-CTCAAACAG------GTACGACAGTC--GTGAAAATTCTCGTTGATGAAAATATGCCTTACGCCCGC Stm TTAACTCTCGTT-CTCAAACAG------GTACGACAGTC--GTGAAAATTCTCGTTGATGAAAATATGCCTTACGCCCGC Sen TTAACTCTCGTT-CTCAAACAG------GTACGACAGTC--GTGAAAATTCTCGTTGATGAAAATATGCCTTACGCCCGC Eco TTAACTCTCGT--CTCATACAG------GTAACACAAAC--GTGAAAATCCTTGTTGATGAAAATATGCCTTATGCCCGC Kpn TTAACTCTCGTT-CTCAGACAG------GTACTGAACT---GTGAAAATCCTCGTTGATGAAAATATGCCCTATGCCCGT Ype CTGTTTTTCATCTGTATGGCAGTTCGCTGTCGGAGAGTAAAGTGAAAATTCTGGTTGATGAAAATATGCCGTACGCTGAG * * ** * * *** ** * ******** ** ***************** ** ** Гены консервативнее, чем межгенные области (точнее, особенности эволюции другие)
 Sty TCGCTCG--CAGCGGAAAGAGGATTACGCCCTTCGCCTGGAGGCTGTGCAGGGGC---GCCGGAGATGGGATGCATAATT Stm TCGCTCG--CAGCGGAAAGAGGATTACGCCCTTCGCCTGGAGGCTGTGCAGGGGC---GCCGGAGATGGGATGCATAATT Sen TCGCTCG--CAGCG")
16
rbsD в E. coli Eco AGGATTAAACTGTGGGTCAGCGAAACGTTTCGCTGATGGAGAAAAAAATGAAAAAAGGC Eco ACCGTTCTTAATTCTGATATTTCATCGGTGATCTCCCGTCTGGGACATACCGATACGCTG

17
rbsD в энтеробактериях Sty AGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Sen AGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Stm GGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Eco AGGATTAAACTGTGGGTCAGCGAAACGTTTCGCTGATGGAGAA-AAAAATGAAAAAAGGC Ype TTTTCTAAACTCCTTGTTAGCGAAACGTTTCGCTCTTGGAGTA-GATCATGAAAAAAGGT ** *** **************** ***** * * ***** ***** Sty ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Sen ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Stm ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Eco ACCGTTCTTAATTCTGATATTTCATCGGTGATCTCCCGTCTGGGACATACCGATACGCTG Ype GTATTACTGAACGCTGATATTTCCGCGGTTATCTCCCGTCTGGGCCATACCGATCAGATT * ** ** **** ** ** **** ** *********** ***** *** *

18
rbsD в энтеробактериях: ответ Sty AGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Sen AGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Stm GGGGTTACACTGCGGC-CAGCGAAACGTTTCGCTAGTGGAGCAGAAAAATGAAGAAAGGC Eco AGGATTAAACTGTGGGTCAGCGAAACGTTTCGCTGATGGAGAA-AAAAATGAAAAAAGGC Ype TTTTCTAAACTCCTTGTTAGCGAAACGTTTCGCTCTTGGAGTA-GATCATGAAAAAAGGT ** *** **************** ***** * * ***** ***** Sty ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Sen ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Stm ACCGTACTCAACTCTGAAATCTCGTCGGTCATTTCCCGTCTGGGGCATACTGATACTCTG Eco ACCGTTCTTAATTCTGATATTTCATCGGTGATCTCCCGTCTGGGACATACCGATACGCTG Ype GTATTACTGAACGCTGATATTTCCGCGGTTATCTCCCGTCTGGGCCATACCGATCAGATT * ** ** **** ** ** **** ** *********** ***** *** *

19
Мораль Комплексный подход: использование многих разнородных соображений, каждое из которых по отдельности – слабое Сравнительный подход: одновременный анализ множества геномов (находящихся на различных эволюционных расстояниях друг от друга)
")
20
Идеология Сходство => гомология (общность происхождения) Гомология => сходная функция Принцип Пирсона: консервативно то, что важно –функциональные мотивы в белках –регуляторные сайты в ДНК –не обязательно последовательности структура белка и РНК расположение генов на хромосоме ко-экспрессия генов
 Гомология => сходная функция Принцип Пирсона: консервативно то, что важно –функциональные мотивы в белках –регуляторные сайты в ДНК –не обязательно последовательности структура белка и РНК расп")
21
Transporters Two main classes –ATP-dependent TM-protein (permease) ATPase Substrate-binding protein –Secondary (symporters, antiporters) Difficult to study in experiment (compared to enzymes) Relatively easy to identify –Similarity to known transporters –Prediction of transmembrane segments Difficult to predict specificity H+H+
 ATPase Substrate-binding protein –Secondary (symporters, antiporters) Difficult to study in experiment (compared to enzymes) Relatively easy to identify –Similarity to known transport")
22
Функциональный анализ транспортеров Предсказание общей функции –гомология –анализ трансмембранных сегментов Отнесение к функциональной подсистеме (метаболическому пути) –ко-локализация –ко-регуляция Предсказание специфичности –анализ филогенетического профиля конечный продукт биосинтетического пути: присутствует в геномах, не имеющих пути (импорт заменяет биосинтез) промежуточный продукт биосинтетического пути; может заменять часть пути «выше по течению» исходный продукт катаболического или биосинтетического пути: не встречается в геномах, где этот путь отсутствует
 –ко-локализация –ко-регуляция Предсказание специфичности –анализ филогенетическог")
23
It is difficult to predict specificity by sequence analysis (nickel-oligopeptide family, substrate-binding NikA)
")
24
PnuC family of cofactor transporters

25
Riboflavin biosynthesis pathway

26
5 UTR regions of riboflavin genes from various bacteria

27
Conserved secondary structure of the RFN-element Capitals: invariant (absolutely conserved) positions. Lower case letters: strongly conserved positions. Dashes and stars: obligatory and facultative base pairs N: any nucleotide. X: any nucleotide or deletion
 positions. Lower case letters: strongly conserved positions. Dashes and stars: obligatory and facultative base pairs N: any nucleotide. X: any nucleotide or d")
28
Attenuation of transcription Terminator The RFN element Antiterminator

29
Attenuation of translation SD-sequestor The RFN element Antisequestor

30
Рибопереключатель RFN: регуляторный механизм Transcription attenuation Translation attenuation

31
YpaA/RibU: транспортёр рибофлавина 5 предсказанных ТМ-сегментов => потенциальный транспортёр регуляторный RFN-элемент => ко- регуляция с генами метаболизма рибофлавина => транспорт рибофлавина или предшественника S. pyogenes, E. faecalis, Listeria: есть ypaA, нет генов биосинтеза рибофлавина => транспорт рибофлавина Предсказание: YpaA – рибофлавиновый транспортёр (Gelfand et al., 1999) Проверка: генетический анализ ( Кренева и др., 2000) биохимический эксперимент ( Burgess et al., 2006)

32
Биотиновый транспортер BioY

33
Метаболическая реконструкция тиаминового биосинтеза = thiN (confirmed) (Gram-positive bacteria) (Gram-negative bacteria) Transport of HMP Transport of HET
 (Gram-positive bacteria) (Gram-negative bacteria) Transport of HMP Transport of HET")
34
yuaJ(=thiT): тиаминовый транспортер (возможно, H + - зависимый) в фирмикутах 6 предсказанных трансмембранных сегментов Почти всегда регулируется THI-рибопереключателями Встречается в геномах, в которых отсутствует тиаминовый путь (Streptococci); В B. cereus импорт тиамина сопряжен с током протонов (Arch. Microbiol., 1977)
: тиаминовый транспортер (возможно, H + - зависимый) в фирмикутах 6 предсказанных трансмембранных сегментов Почти всегда регулируется THI-рибопереключателями Встречается в геномах, в которых отсутствует тиаминовый путь (Streptococci); В B.")
35
Почти всегда регулируются THI-рибопереключателями Не встречаются в геномах, в которых отсутствует тиаминовый путь Всегда встречаются вместе с thiD и thiE В ряде геномов (Pasteurellacee, Brucella некоторые фирмикуты) встречаются в отсутствие thiC thiX-thiY-thiZ и ykoF-ykoE-ykoD- ykoC: предсказанные АТФ-зависимые транспортеры HMP
 встречаются в отсутствие thiC thiX")
36
Co и Ni ко-локализация (хромосомные локусы) –транспортеры Ni – с генами никель-зависимых ферментов –транспортеры Co – с генами синтеза кобаламина ко-регуляция –транспортеры Ni – фактор транскрипции NikR –транспортеры Co – рибопереключатель В12
 –транспортеры Ni – с генами никель-зависимых ферментов –транспортеры Co – с генами синтеза кобаламина ко-регуляция –транспортеры Ni – фактор транскрипции NikR –транспортеры Co – рибопереключатель В12")
37
Пять семейств транспортеров

38
Новое семейство транспортеров Co и Ni NikM CbiM Ni 2+ Co 2+ + CbiN + NikL, NikK + NikN + NikL

39
Структура локусов B12-элементсайт связывания NikRгены

40
Структура cbiO=NikO~bioM cbiQ=NikQ~bioN

41
Проверка: тест на транспорт ионов Co Ni

42
Для транспорта достаточно компонент МN (первый пример такого АВС-транспортера) cbiMNQO cbiMNQ cbiMN cbiM контроль
 cbiMNQO cbiMNQ cbiMN cbiM контроль")
43
Вспомним BioY. Действительно, BioY достаточно; у BioMNY более крутая кинетика

46
Экспериментальные подтверждения рибофлавинтиаминфолат

47
Дмитрий Родионов –регуляция транскрипции –метаболическая реконструкция –идентификация транспортеров Алексей Витрещак –рибопереключатели А.А. Миронов –программное обеспечение Томас Хеббельн (Берлин) – Cо, Ni, биотин Андрей Остерман (Сан Диего) – рибофлавин Эндрю Хансон (Флорида) – тиамин Дирк Слотблум (Гронинген) – фолат
 – Cо, Ni, биотин Андрей Остерман (Сан Диего) – рибоф")
49
Transporters Two main classes –ATP-dependent TM-protein (permease) ATPase Substrate-binding protein –Secondary (symporters, antiporters) Difficult to study in experiment (compared to enzymes) Relatively easy to identify –Similarity to known transporters –Prediction of transmembrane segments Difficult to predict specificity H+H+
 ATPase Substrate-binding protein –Secondary (symporters, antiporters) Difficult to study in experiment (compared to enzymes) Relatively easy to identify –Similarity to known transport")
50
It is difficult to predict specificity by sequence analysis (nickel-oligopeptide family, substrate-binding NikA)
")
51
PnuC family of cofactor transporters

52
Riboflavin biosynthesis pathway

53
5 UTR regions of riboflavin genes

54
RFN-element Capitals: invariant (absolutely conserved) positions. Lower case letters: strongly conserved positions. Dashes and stars: obligatory and facultative base pairs Degenerate positions: R = A or G; Y = C or U; K = G or U; B= not A; V = not U. N: any nucleotide. X: any nucleotide or deletion
 positions. Lower case letters: strongly conserved positions. Dashes and stars: obligatory and facultative base pairs Degenerate positions: R = A or G; Y = C or U; K = G or U; B= not A; V = not U.")
55
RFN: the mechanism of regulation Transcription attenuation Translation attenuation

56
YpaA: riboflavin transporter 5 predicted TM segments => a transporter Upstream RFN element => co-regulation with riboflavin genes => transport of riboflavin / precursor S. pyogenes, E. faecalis, Listeria spp.: ypaA, no riboflavin pathway => transport of riboflavin Prediction: YpaA is riboflavin transporter (Gelfand et al., 1999) Verification: by genetic analysis (Kreneva et al., 2000) directly (Burgess et al., 2006) => RibU ypaA is regulated by riboflavin (Lee et al., 2001) … via attenuation of transcription (Winkler et al., 2003)

57
Biotin transporter BioY Identification: –co-localization –co-regulation –phylogenetic profiling Additional components –ATPase(?) bioM –Permease(?) bioN
 bioM –Permease(?) bioN")
58
Thiamin biosynthesis = thiN (confirmed) (Gram-positive bacteria) (Gram-negative bacteria) Transport of HMP Transport of HET
 (Gram-positive bacteria) (Gram-negative bacteria) Transport of HMP Transport of HET")
59
yuaJ(=thiT): thiamine transporter 6 predicted TM-segments Regulated by THI riboswitches Streptococci: ThiT, no thiamine pathway
: thiamine transporter 6 predicted TM-segments Regulated by THI riboswitches Streptococci: ThiT, no thiamine pathway")
60
Regulated by THI riboswitches Newer occurs in genomes lacking thiamine pathway Always co-occurs with thiD and thiE Sometimes occurs without thiC ykoFEDC: ATP-dependent HMP transporter

61
Cobalt and Nickel Co-localization –Ni transporters with genes for Ni- dependent enzymes –Co transporters with cobalamine biosynthesis genes Co-regulation –Ni transporters by transcription factor NikR –Co transporters by В12 riboswitich

62
Structure of the loci B12 riboswitchNikR binding sitegenes

63
Five families of transporters

64
New ATP-dependent transporters NikM CbiM Ni 2+ Co 2+ + CbiN + NikL, NikK + NikN + NikL

65
Dmitry Rodionov Thomas Eitinger

66
Test 1: predicted specificity is correct Co Ni

67
Structure: too many components

68
Biotin transporter BioY ATPase BioM ~ CbiO = NikO Permease BioN ~ CbioQ = NikQ

69
Test 2: MN components are suffucient (ATPase and permease are dispensable) cbiMNQO cbiMNQ cbiMN cbiM control
 cbiMNQO cbiMNQ cbiMN cbiM control")
70
Test 3: BioY is sufficient Even if the genome had BioMNY; BioMNY has better cinetics

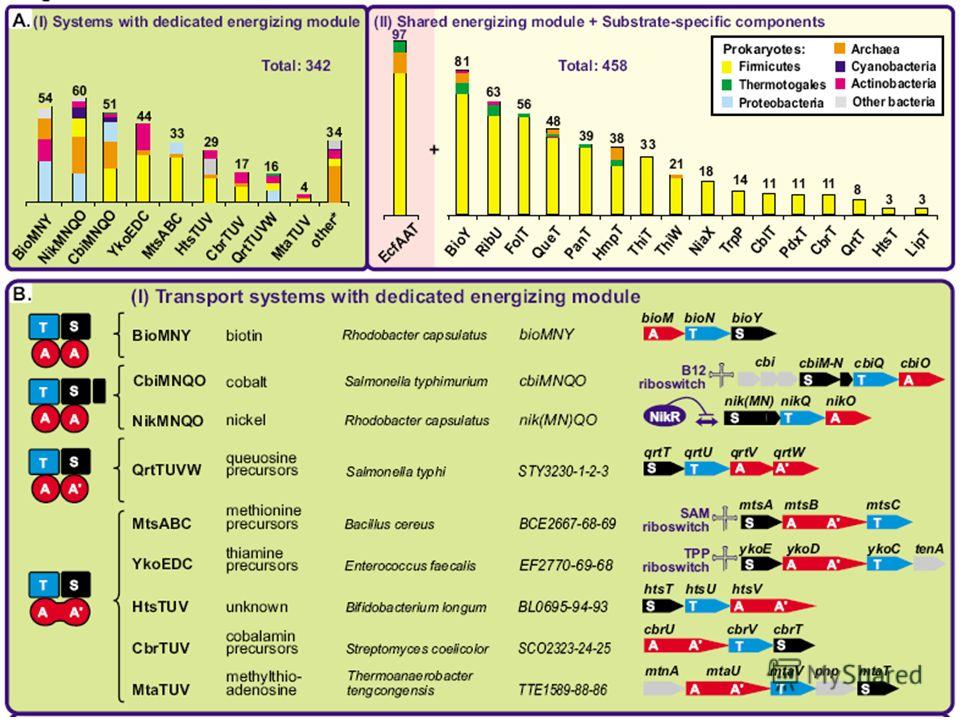
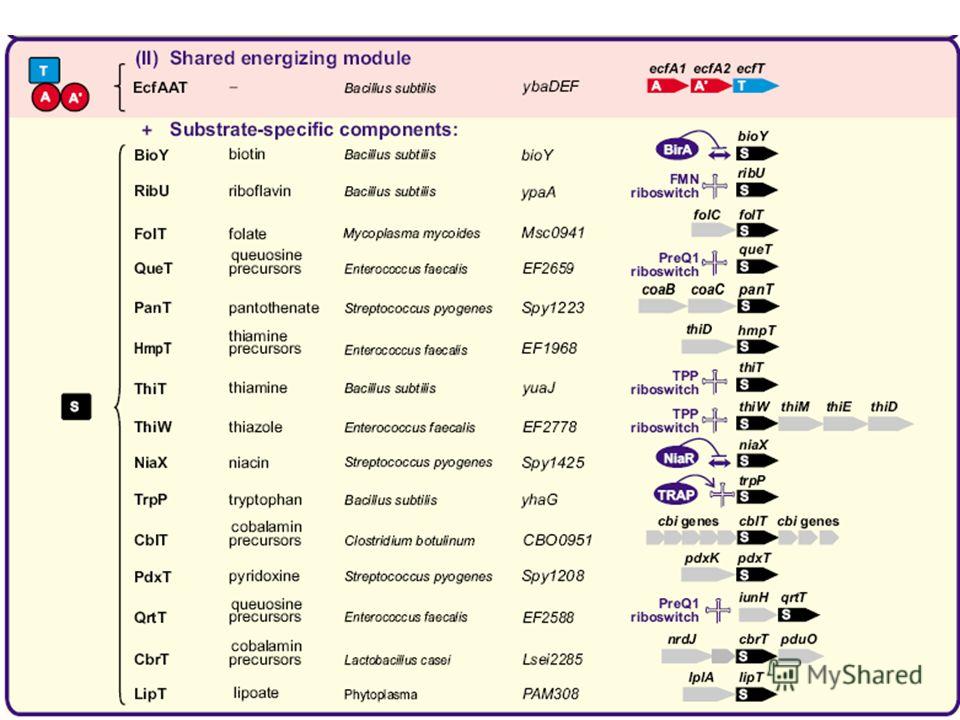
71
Tip of the iceberg?

75
Validations RibU: riboflavinThiT: thiamin FolT: folate (like BioY)
")
76
Universal energizing component + specific components

77
Цель (глобальная) Предсказать свойства организма путем (компьютерного) анализа его генома (возможно, с использованием дополнительной информации: эпигенетика, белок-белковые взаимодействия и т.п.) сейчас: метаболическая реконструкция, транспортные системы, ответ на стресс и т.д. Понять эволюцию геномов/организмов
 Предсказать свойства организма путем (компьютерного) анализа его генома (возможно, с использованием дополнительной информации: эпигенетика, белок-белковые взаимодействия и т.п.) сейчас: метаболическая реконструкция, транспортные сис")
78
«Неприкладная» биоинформатика Молекулярная эволюция –филогения генов –таксономия организмов –горизонтальные переносы и т.п. –положительный и отрицательный отбор что сделало нас людьми? лекарственная устойчивость –эволюция геномов Системная биология –строение геномов –сети взаимодействий белок-белковые регуляция транскрипции сигнальные пути

79
Задачи биоинформатики С проверяемым ответом –предсказание функции, регуляции, структуры и т.п.: ставим эксперимент С непроверяемым ответом –эволюционные деревья но если бы знать все геномы всех (в том числе очень давно умерших) существ, то задача станет тривиальной С принципиально непроверяемым ответом (который зависит от операциональных определений) –идентификация повторов, консервативных областей, островов метилирования и т.п. (так ли он непроверяем?) Без ответа (общеописательные) –статистика геномов (изохоры и т.п.) –описание регуляторных и пр. сетей (hubs, мотивы и т.п.)
 существ, то задача ста")
80
«В принципе не проверяемые ответы» (зависящие от определений) Так ли они непроверяемы? Повторы –если иметь все геномы, то можно описывать вставки/замены фрагментов генома и их последующее расхождение Консервативные области –если иметь все геномы, то можно просто оценивать локальную скорость эволюции (но это будет функцией времени) Статистика ДНК (локальный нуклеотидный состав) –это следствие локального паттерна замен, так и надо описывать Микросателлиты –можно ли «функционально» (а не операционально) определить микросателлит, исходя из динамики вставок/замен/дупликаций? CpG-острова –можно ли «функционально» (а не операционально) определить CpG-остров, исходя из паттерна мутаций, состояния метилирования и т.п.? (тут уже эволюция + эксперимент)
 Так ли они непроверяемы? Повторы –если иметь все геномы, то можно описывать вставки/замены фрагментов генома и их последующее расхождение Консервативные области –если иметь все геномы, то")
81
Цель (недостижимая?) откуда оно все взялось? первое приближение – реконструкция генома/свойств LUCA реально ли заглянуть глубже? реально ли смоделировать? (времена) реально ли смоделировать «по частям»?
 откуда оно все взялось? первое приближение – реконструкция генома/свойств LUCA реально ли заглянуть глубже? реально ли смоделировать? (времена) реально ли смоделировать «по частям»?")
Еще похожие презентации в нашем архиве:




 М.С.Гельфанд 23 января 2009.")

 А. Неверов (задания) Е. Ермакова,")

 Е.Ермакова (задания, занятия)")
 Д. Равчеев (задания)")


 молекулярной биологии – матричный синтез. Этапы биосинтеза белка: ДНК репликация ДНК транскрипция.")





 Q: Что было бы для двух других моделей? Коетки растут.")





