Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Катализ Возможность протекания хим.реакций обусловлена разницей свободной энергии исх. веществ и продуктов. Самопроизвольное течение реакции возможно, если суммарная энергия (G) исходных веществ выше, чем продуктов (реакция экзергоническая). Реакция невозможна, если G ниже (реакция эндергоническая). Скорость экзергонической реакции зависит от «энергетического барьера», который нужно преодолеть веществом. У реакционно способных молекул энергии достаточно для преодоления барьера. Энергетическая активация – дополнительное количество энергии, необходимое молекулярным веществам для преодоления энергетического барьера. Катализ – ускорение хим.реакций веществами участвующих в процессе, но не расходующихся. Катализаторы – вещества ускоряющие химические реакции. Катализ – ускорение хим.реакций веществами участвующих в процессе, но не расходующихся. Катализаторы – вещества ускоряющие химические реакции. Ферменты снижают энергию активации (Еа) (понижают энергетический барьер) процесса, воздействуя на хим. связи реагируемых веществ. В результате возрастает доля активированных молекул, возрастает скорость реакции. Ферменты снижают энергию активации (Еа) (понижают энергетический барьер) процесса, воздействуя на хим. связи реагируемых веществ. В результате возрастает доля активированных молекул, возрастает скорость реакции.
 исходных веществ выше, чем продуктов (реакция экзергоническая). Реакция")
2
Изменение свободной энергии при разложении угольной кислоты.

3
Изменение свободной энергии в ходе химической реакции, некатализируемой и катализируемой ферментами. Фермент понижает энергию активации Еа, т.е. снижает высоту энергетического барьера, в результате возрастает доля реакционно- способных молекул, следовательно, увеличивается скорость реакции. Изменение свободной энергии.

4
АКТИВНОСТЬ ФЕРМЕНТОВ АКТИВНОСТЬ ФЕРМЕНТОВ Активность ферментов определяют косвенно: 1. по количеству образующегося продукта (Р), 2. по количеству потребляемого субстрата (S). Международная единица активности фермента или Е – количество фермента, превращающее 1 мкмоль S в 1 мин. в стандартных условиях в расчете на 1 г ткани. Удельная активность – количество Е / мг белка.
, 2. по количеству потребляемого субстрата (S). Международная единица активности фермента или Е – количество фермента, прев")
5
Для оценки количества молекул фермента среди других белков данной ткани определяют удельную активность фермента, численно равную количеству превращенного субстрата (мкмоль), делённому на массу (мг) белка в этой ткани в единицу времени (мин): Активность ферментов.
, делённому на массу (мг) белка в этой ткани в единицу времени (мин): Активнос")
6
Каталитическая эффективность. реакций высокоэффективны, они протекают в раз быстрее, чем не катализируемые реакции. Большинство катализируемых ферментами реакций высокоэффективны, они протекают в раз быстрее, чем не катализируемые реакции. Каждая молекула фермента способна за секунду трансформировать от 100 до 1000 молекул субстрата в продукт. Количество молекул субстрата, превращенных в продукт с помощью одной молекулы фермента за 1 с, называют, или молярной активностью. Количество молекул субстрата, превращенных в продукт с помощью одной молекулы фермента за 1 с, называют числом оборотов фермента, или молярной активностью.

7
Специфичность. Специфичность наиболее важное свойство ферментов, определяющее биологическую значимость этих молекул. Различают субстратную и каталитическую специфичности фермента, определяемые строением активного центра.

8
Субстратная специфичность ферментов В основе лежит строгое соответствие размеров и структуры субстратов активному центру. 1. Абсолютная специфичность – действие только на один субстрат (уреаза взаимодействует только с мочевиной). 2. Разновидность – стехиометрическая (оптическая) специфичность – действие только на определенные стереоизомеры. Фумараза действует только на фумаровую кислоту. 3. Относительная (групповая) – действие на группу веществ имеющих один тип связи. Амилаза – на крахмал, гликоген, декстрины, пепсин – на разные белки.
. 2. Разновидность – с")
9
Каталитическая специфичность Биологическая функция фермента, как и любого белка, обусловлена наличием в его структуре активного центра. Лиганд, взаимодействующий с активным центром фермента, называют субстратом. В активном центре фермента есть аминокислотные остатки, функциональные группы которых обеспечивают связывание субстрата, и аминокислотные остатки, функциональные группы которых осуществляют химическое превращение субстрата. Условно эти группы обозначают как участок связывания субстрата и каталитический участок, однако не всегда эти участки имеют чёткое пространственное разделение и иногда могут «перекрываться».

10
Полиферментные системы 1. Каждая клетка имеет специфичный состав ферментов. 2. Некоторые ферменты содержатся во всех клетках, другие в немногих. 3. Работа каждого фермента, обычно, не индивидуальна, а связана с другими ферментами из которых формируется полиферментные системы – конвейеры. 4. Субстрат проходит длинную цепь реакции многих ферментов Р 1S 2 Р 2S 3 5. Некоторые ферменты системы связаны с органеллами, биомембранной или цитоскелетом. 6. Некоторые ферменты одной цепи метаболизма объединяются в мультиферментные комплексы с определенной функцией.

11
Ферментные ансамбли или мультиферменты – комплекс ферментов, катализирующих последовательные реакции при превращении одного вещества.

12
Энергетические изменения ПРИ ХИМИЧЕСКИХ РЕАКЦИЯХ. Любые химические реакции протекают, подчиняясь двум основным законам термодинамики (сохранения энергии и энтропии). Согласно этим законам, общая энергия химической системы и её окружения остаётся постоянной, при этом химическая система стремится к снижению упорядоченности (увеличению энтропии).
. Согласно этим законам, общая энергия химической системы и её окружения остаётся пост")
13
Изменение свободной энергии в ходе химической реакции, некатализируемой и катализируемой ферментами. Фермент понижает энергию активации Еа, т.е. снижает высоту энергетического барьера, в результате возрастает доля реакционно-способных молекул, следовательно, увеличивается скорость реакции. Изменение свободной энергии.

15
Механизм ферментативного катализа. В механизме ферментативного катализа решающее значение имеет образование нестойких промежуточных соединений фермент- субстратный комплексов – [ES], подвергающихся превращению в нестабильный переходной комплекс – [ЕР], который почти мгновенно распадается на свободный фермент и продукт реакции.
![Механизм ферментативного катализа. В механизме ферментативного катализа решающее значение имеет образование нестойких промежуточных соединений фермент- субстратный комплексов – [ES], подвергающихся превращению в нестабильный переходной комплекс – [ЕР](http://images.myshared.ru/6/529940/slide_15.jpg "Механизм ферментативного катализа. В механизме ферментативного катализа решающее значение имеет образование нестойких промежуточных соединений фермент- субстратный комплексов – [ES], подвергающихся превращению в нестабильный переходной комплекс – [ЕР")
16
Образование фермент-субстратного комплекса. где Е фермент (энзим), S субстрат, Р продукт. Данные обозначения общеприняты и происходят от английских слов enzyme, substrat, produkt. Схематично процесс катализа можно представить следующим уравнением:
, S субстрат, Р продукт. Данные обозначения общеприняты и происходят от английских слов enzyme, substrat, produkt. Схематично процесс катализа можно представить следующим уравнением:")
18
Эффекты деформации и индуцированного соответствия. Развитие теории Э.Фишера «ключ - замок»: Активный центр фермента также способствует дестабилизации межатомных связей в молекуле субстрата, что облегчает протекание химической реакции и образование продуктов. Это свойство активного центра называют эффектом деформации субстрата. Свойство активного центра «подстраиваться под субстрат» названо теорией индуцированного соответствия (Д. Кошленд).

19
В участке связывания субстрат при помощи нековалентных связей взаимодействует (связывается) с ферментом, формируя фермент- субстратный комплекс. В участке связывания субстрат при помощи нековалентных связей взаимодействует (связывается) с ферментом, формируя фермент- субстратный комплекс. В каталитическом участке субстрат претерпевает химическое превращение в продукт, который затем высвобождается из активного центра фермента. Последовательность действия ферментов
 с ферментом, формируя фермент- субстратный комплекс. В участке связывания субстрат при помощи нековалентных связей взаимодействует (связывается) с ферментом,")
20
Механизм действия ферментов 1. Стадия: диффузия, связывание S ферментом Е и образование фермент – субстратного комплекса - ES. Индуцированное соответствие S активному центру. Еа измеряется незначительно. Очень быстрый процесс. 2. Стадия: образование активированного ES*. Резкое снижение Еа. Взаимодействие Е и S. Дестабилизация химических связей в S. 3. Стадия: образование продуктов и их выход.

23
Зависимость скорости реакции (V) от концентрации субстрата S. Vmax максимальная скорость реакции при данной концентрации фермента в оптимальных условиях проведения реакции. Кm константа Михаэписа.
 от концентрации субстрата S. Vmax максимальная скорость реакции при данной концентрации фермента в оптимальных условиях проведения реакции. Кm константа Михаэписа.")
24
Мультисубстратные реакции: Большинство ферментов катализирует реакции, в которых участвует более чем один субстрат. В случае если кофермент не является простетической группой, его также можно рассматривать как ещё один субстрат. Следовательно, участников ферментативной реакции может быть несколько: непосредственно фермент, несколько субстратов и кофермент.

25
Механизм упорядоченного взаимодействия субстрата с активным центром фермента: Первым в активный центр фермента присоединяется субстрат А, облегчая присоединение субстрата В. После химической модификации также наблюдают определенный порядок высвобождения продуктов реакции.

26
Этапы ферментативного катализа. I - этап сближения и ориентации субстрата относительно активного центра фермента; II - образование фермент- субстратного комплекса (ES) в результате индуцированного соответствия; III - деформация субстрата и образование нестабильного комплекса фермент-продукт (ЕР); IV - распад комплекса (ЕР) с высвобождением продуктов реакции из активного центра фермента и освобождением фермента. I - этап сближения и ориентации субстрата относительно активного центра фермента; II - образование фермент- субстратного комплекса (ES) в результате индуцированного соответствия; III - деформация субстрата и образование нестабильного комплекса фермент-продукт (ЕР); IV - распад комплекса (ЕР) с высвобождением продуктов реакции из активного центра фермента и освобождением фермента.
 в результате индуцированного соответствия; III - деформация субстрата и образовани")
27
Молекулярные механизмы ферментативного катализа 1. Эффект ориентации реагентов снижает энтропию и энергию активации, ускоряет реакцию в тысячи раз. 2. Эффект «деформации» субстрата – «растягивается» хим.связь, снижается энергия её разрыва (снижается энергия активации). 3. Кислотно-основной катализ. В активном центре имеются функциональные группы аминокислотных остатков с кислотными и основными группами. Фермент является и акцептором и донором протонов и электронов. Происходит перераспределение электронной плотности на участке субстрата. Это облегчает перестройку и разрыв связей. 4. Ковалентный катализ – образование ковалентных связей с субстратами.

28
Снижение энергетического барьера происходит за счет: 1. Повышения вероятности столкновения субстратов. 2. Строгая ориентация взаимодействия молекул в активном центре. 3. Максимальное сближение субстратов. 4. Действие на определённые атомы субстрата атомами активного центра. 5. Смещение электронов и протонов, что повышает реакционоспособность атомов.

29
Ферментативная кинетика Раздел биохимии, который изучает ферментативную активность в зависимости от условий протекания реакций. В частности, в зависимости от количества субстратов, продуктов и других метаболитов. В зависимости от температуры, давления, кислотности среды и наличия ингибиторов.

30
Зависимость скорости ферментативной реакции (V) от концентрации фермента.
 от концентрации фермента.")
31
Зависимость скорости ферментативной реакции (V) от температуры.
 от температуры.")
32
Зависимость скорости ферментативной реакции (V) от рН среды.
 от рН среды.")
33
Оптимальные значения рН для некоторых ферментов.

34
Зависимость скорости реакции (V) от концентрации субстрата S. Vmax максимальная скорость реакции при данной концентрации фермента в оптимальных условиях проведения реакции. Кm константа Михаэписа.
 от концентрации субстрата S. Vmax максимальная скорость реакции при данной концентрации фермента в оптимальных условиях проведения реакции. Кm константа Михаэписа.")
35
Схема конкурентного ингибирования активности фермента.

38
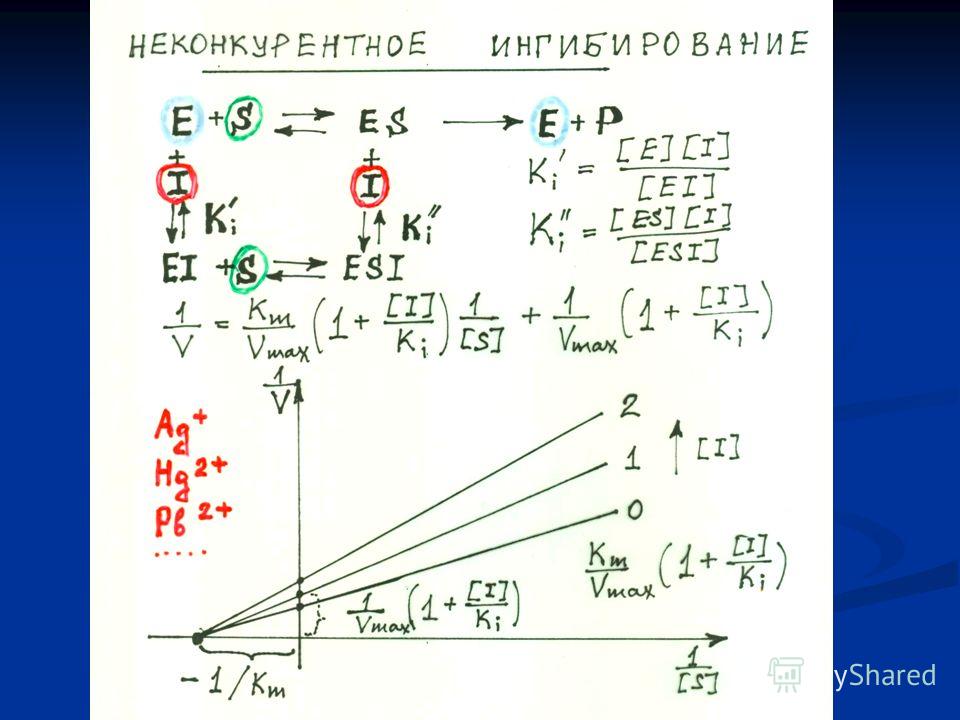
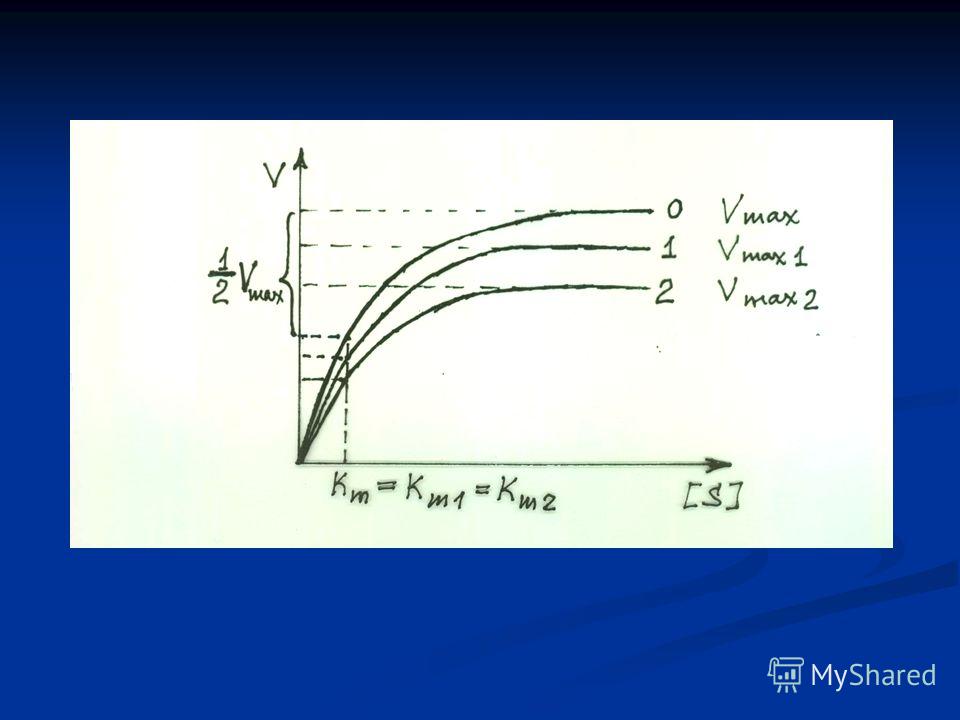
Схема неконкурентного ингибирования активности фермента.

41
Механизм действия ионов ртути как необратимого ингибитора. Ионы ртути в малых концентрациях блокируют сульфгидрильные группы активного центра, что приводит к снижению скорости ферментативной реакции.

42
Основные способы регуляции активности ферментов: аллостерическая регуляция; аллостерическая регуляция; регуляция с помощью белок-белковых взаимодействий; регуляция с помощью белок-белковых взаимодействий; регуляция путём фосфорилирования/дефос- форилирования молекулы фермента; регуляция путём фосфорилирования/дефос- форилирования молекулы фермента; регуляция частичным (ограниченным) протеолизом. регуляция частичным (ограниченным) протеолизом.

43
Схема положительной и отрицательной регуляции катаболизма глюкозы. Молекула АТФ участвует в ретроингибировании аллостерических ферментов фосфо-фруктокиназы и пируваткиназы. Фруктозо- 1,6-бисфосфат активатор метаболического пути распада глюкозы. Плюсами отмечена активация, минусами ингибирование ферментов.

44
Регуляция активности ферментов фосфорилированием/дефосфорилиро ванием.

45
Изоформы лактатдегидрогеназы. А строение различных изоформ ЛДГ; Б распредепение на электрофореграмме и относительные количества изоформ ЛДГ в различных органах; В содержание изоформ ЛДГ в плазме крови в норме и при патологии (электрофореграммы слева и фотометрическое сканирование справа).

46
Изменение активности ферментов в плазме крови при инфаркте миокарда.

47
Лабильность ферментов Каталитическая эффективность фермента, как и любой белковой молекулы, зависит от его конформации, и в частности от конформации активного центра. Для ферментов характерна конформационная лабильность способность к небольшим изменениям нативной конформации вследствие разрыва слабых связей. Поэтому воздействие денатурирующих агентов, способных изменять конформацию молекулы фермента, приводит к изменению конформации активного центра и снижению способности присоединять субстрат. В результате этого уменьшается каталитическая эффективность фермента.

48
Возможные пути преобразования глюкозы-6-фосфата.

Еще похожие презентации в нашем архиве:






 - это высокоспецифичные белки, выполняющие функции биологических катализаторов. Катализатор - это вещество, которое ускоряет.")
 Совокупность протекающих в клетке химических превращений, обеспечивающих её рост,")










. Значение параметра.")



. Столкновение частиц Число соударений в газе 10 10 за с при н.у. мгновенное протекание реакции? Кроме факта столкновения,")



