Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
СЛОЖНЫЕ БЕЛКИ

2
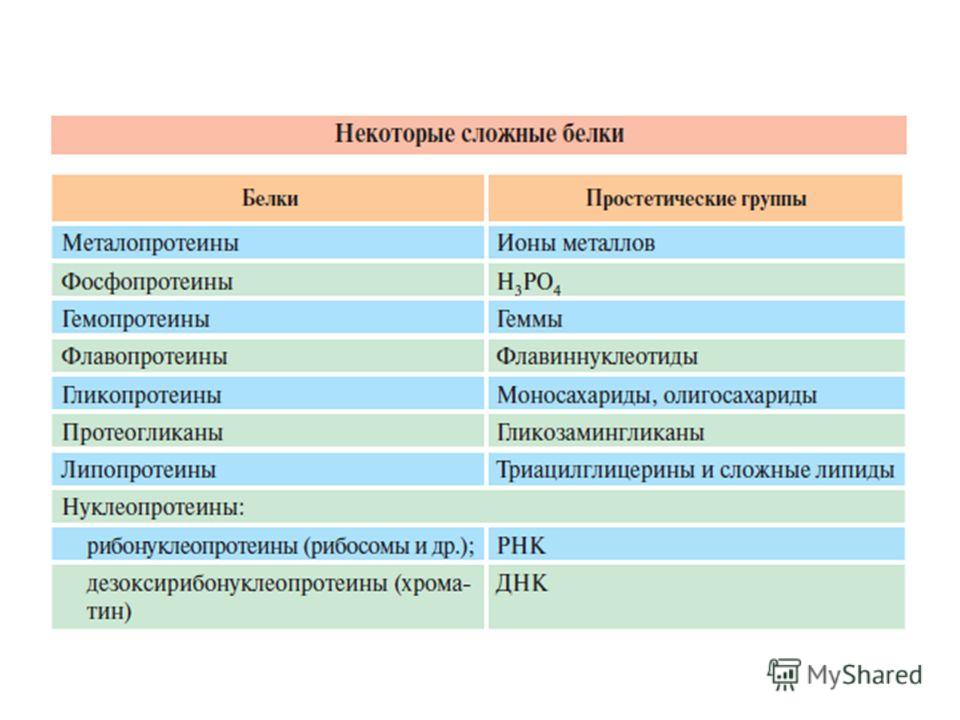
Если белки кроме пептидных цепей содержат еще компоненты неаминокислотной природы, то такие белки называются сложными. Небелковую часть называют простетической группой, а белковую апопротеином. Сложный белок холопротеин может диссоциировать на компоненты: холопротеин апопротеин + простетическая группа. Направление реакции зависит от прочности связи компонентов холопротеина. Простетической группой могут быть органические вещества, ионы металлов, нуклеиновые кислоты, углеводы, липиды и другие вещества

4
Функции простетической группы в составе сложного белка разнообразны: - изменяет свойства белков (заряд, растворимость, термолабильность), например, фосфорная кислота в фосфопротеинах или остатки моносахаридов в гликопротеинах, - защищает белок от протеолиза вне и внутри клетки, например углеводная часть в гликопротеинах, - в виде лиганда обеспечивается транспорт нерастворимых в воде соединений, например, перенос жиров липопротеинами,
, например, фосфорная кислота в фосфопротеинах или остатки моносахаридов в гликопротеинах, - защищает белок от про")
5
- придает биологическую активность и определяет функцию белка, например, нуклеиновая кислота в нуклеопротеинах, гем в гемоглобине, углевод в рецепторных белках, - влияет на проникновение через мембраны, внутриклеточную миграцию, сортировку и секрецию белков. Это выполняет, как правило, углеводный остаток.

6
Хромопротеины (от греч. chroma – краска) состоят из простого белка и связанного с ним окрашенного небелкового компонента. Различают гемопротеины, магнийпорфирины и флавопротеины (содержат производные изоаллоксазина).
 состоят из простого белка и связанного с ним окрашенного небелкового компонента. Различают гемопротеины, магнийпорфирины и флавопротеины (содержат производные изоаллоксазина).")
7
Гемопротеины: - неферментативные (гемоглобин, миоглобин) -ферменты (цитохромы, каталаза, пероксидаза).
 -ферменты (цитохромы, каталаза, пероксидаза).")
8
Гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями. Это - небелковая часть гемопротеинов
 и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями. Это - небелковая часть гемопротеинов")
9
Все 4 связи с атомами азота пиррольных колец расположены в одной плоскости, 5-я и 6-я координационные связи (с атомом азота имидазольного кольца гистидина и с кислородом соответственно) – по разные стороны перпендикулярно к этой плоскости
 – по разные стороны перпендикулярно к этой плоскости")
11
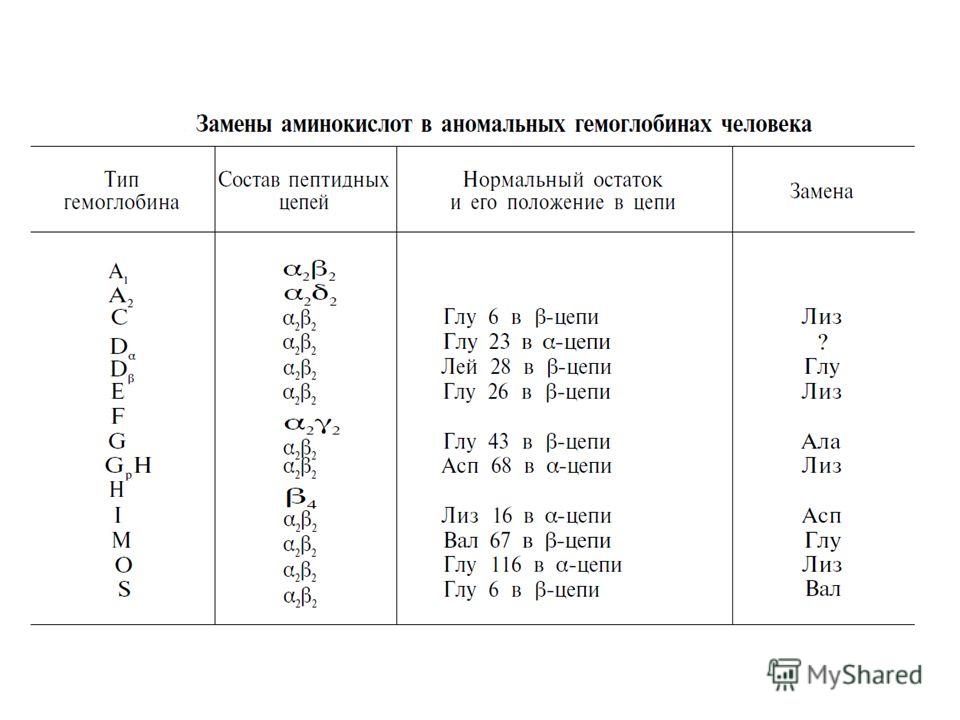
Болезни гемоглобинов (их насчитывают более 200) называют гемоглобинозами. Принято делить их на гемоглобинопатии, в основе развития которых лежит наследственное изменение структуры какой-либо цепи нормального гемоглобина (часто их относят также к молекулярным болезням ), и талассемии, обусловленные наследственным нарушением синтеза какой-либо нормальной цепи гемоглобина. Различают также железодефицитные анемии.
 называют гемоглобинозами. Принято делить их на гемоглобинопатии, в основе развития которых лежит наследственное изменение структуры какой-либо цепи нормального гемоглобина (часто их относят также к моле")
12
Флавопротеины Флавопротеины содержат прочно связанные с белком простетические группы, представленные изоаллоксазиновыми производными – окисленными флавинмононуклеотидом (ФМН) и флавинадениндинуклеотидом (ФАД). Флавопротеины входят в состав оксидоредуктаз – ферментов, катализирующих окислительно- восстановительные реакции в клетке. Типичными представителями флавопротеинов, содержащих также негемовое железо, являются ксантиноксидаза, альдегидоксидаза, СДГ, дигидрооротатдегидрогеназа, ацил-КоА-дегидрогеназа
 и флавинадениндинуклеотидом (ФАД). Флавопротеины входят в состав оксидореду")
13
НУКЛЕОПРОТЕИНЫ Нуклеопротеины состоят из белков и нуклеиновых кислот. Последние рассматриваются как простетические группы. В природе обнаружено 2 типа нуклеопротеинов, отличающихся друг от друга по составу, размерам и физико-химическим свойствам,– дезоксирибонуклеопротеины (ДНП) и рибонуклеопротеины (РНП).

14
ЛИПОПРОТЕИНЫ В последние годы достигнут определенный прогресс в выяснении химической природы и структуры липопротеинов (ЛП). Этот класс сложных белков состоит из белка и простетической группы, представленной каким- либо липидом. В частности, в составе липопротеинов открыты нейтральные жиры, свободные жирные кислоты, фосфолипиды, холестериды.
. Этот класс сложных белков состоит из белка и простетической группы, представленной каким- либо липидом. В частности, в состав")
16
ФОСФОПРОТЕИНЫ К белкам этого класса относятся казеиноген молока, вителлин, вителлинин и фосвитин, выделенные из желтка куриного яйца; овальбумин, открытый в белке куриного яйца; ихтулин, содержащийся в икре рыб, и др. Большое количество фосфопротеинов содержится в клетках ЦНС. В клетках фосфопротеины синтезируются в результате посттрансляционной модификации, подвергаясь фосфорилированию при участии протеинкиназ,, катализирующих фосфорилирование ОН-группы тирозина, серина и треонина.

17
ГЛИКОПРОТЕИНЫ Гликопротеины – сложные белки, содержащие, помимо простого белка или пептида, группу гетероолигосахаридов. В настоящее время их принято называть гликоконъюгатами. В состав гликоконъюгата входит углеводный компонент (гликановая фракция), ковалентно связанный с неуглеводной частью (агликановая фракция), представленной белком, пептидом, аминокислотой или липидом.
,")
18
К типичным гликопротеинам относят большинство белковых гормонов, мембранные сложные белки, антитела (иммуноглобулины), белки плазмы крови, молока, овальбумин, интерфероны, факторы комплемента, группы крови, рецепторные белки. Гликопротеины выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное и гормональное, а также антивирусное действие.
, белки плазмы крови, молока, овальбумин, интерфероны, факторы комплемента, группы крови, рецепторные белки. Гликопротеины выполняют")
19
Типы связей между углеводными компонентами и белками включают О-гликозидные связи (с ОН- группами серина, треонина и оксилизина), N- гликозидные связи (с амидными группами аспарагина, реже глутамина или ε-NH2 группами лизина и аргинина) и эфирные гликозидные связи со свободными СООН-группами глутаминовой и аспарагиновой кислот.
, N- гликозидные связи (с амидными группами аспарагина, реже глутамина или ε-NH2 группами лизина и аргинина) и эфирные гли")
20
МЕТАЛЛОПРОТЕИНЫ К металлопротеинам относятся биополимеры, содержащие, помимо белка, ионы какого-либо одного металла или нескольких металлов. К таким белкам принадлежат, например, белки, содержащие негемовое железо, а также белки, координационно связанные с атомами металлов в составе сложных белков-ферментов. Типичными представителями первых являются железосодержащие белки ферритин, трансферрин и гемосидерин

21
Ко второй группе металлопротеинов относится ряд ферментов: ферменты, содержащие связанные с молекулой белка ионы металлов, определяющих их функцию,– металлоферменты (в процессе очистки металлы остаются связанными с ферментами)
")
22
ФЕРМЕНТЫ

23
Ферментами (энзимами) называют растворимые или связанные с мембранами белки, наделенные каталитической активностью. Кроме белков каталитическую активность в организме могут проявлять некоторые РНК (рибозимы) и антитела (абзимы), однако они в тысячи раз менее эффективны, чем ферменты.)
 называют растворимые или связанные с мембранами белки, наделенные каталитической активностью. Кроме белков каталитическую активность в организме могут проявлять некоторые РНК (рибозимы) и антитела (абзимы), однако они в тысячи р")
24
S - исходный субстрат; Р - продукт; ΔЕНФ -энергия активации неферментативной реакции; ΔЕФ - энергия активации ферментативной реакции; ΔG - стандартное изменение свободной энергии.

25
Фермент снижает энергию активации (энергия, необходимая для перевода всех молекул моля вещества в активированное состояние при данной температуре) путем увеличения числа активированных молекул, которые становятся реакционноспособными на более низком энергетическом уровне. Следует отметить, что как катализируемая ферментом, так и не катализируемая им реакция независимо от ее пути имеет одинаковую величину стандартного изменения свободной энергии (ΔG). Действуя на скорость реакции, ферменты не изменяют равновесия между прямой и обратной реакциями, константа равновесия реакции остается постоянной; они лишь ускоряют наступление равновесия химической реакции
 путем увеличения числа активированных молекул, которые становятся реакционноспособными на более низком")
26
а - реакция первого порядка (при [S] < Кm скорость реакции пропорциональна концентрации субстрата); б - реакция смешанного порядка; в - реакция нулевого порядка, когда v = Vmax и скорость реакции не зависит от концентрации субстрата
![а - реакция первого порядка (при [S] < Кm скорость реакции пропорциональна концентрации субстрата); б - реакция смешанного порядка; в - реакция нулевого порядка, когда v = Vmax и скорость реакции не зависит от концентрации субстрата](http://images.myshared.ru/5/492883/slide_26.jpg "а - реакция первого порядка (при [S] < Кm скорость реакции пропорциональна концентрации субстрата); б - реакция смешанного порядка; в - реакция нулевого порядка, когда v = Vmax и скорость реакции не зависит от концентрации субстрата")
27
Отличительной особенностью ферментов является наличие активного центра. Активный центр - это уникальная комбинация сближенных в пространстве аминокислотных остатков, которая обеспечивает: а) узнавание молекулы субстрата, б) связывание субстрата с ферментом, в) осуществление каталитического превращения (в случае сложного фермента в акте катализа также принимает участие кофермент, входящий в состав активного центра).
 узнавание молекулы субстрата, б) связывание субстрата с фер")
28
Одна молекула фермента, особенно если она состоит из нескольких субъединиц, может содержать более одного активного центра. В активном центре имеются два участка. Первый участок отвечает за узнавание и связывание субстрата. Он называется субстрат-связывающим участком или якорной площадкой. Второй участок называется каталитическим, в его состав входят аминокислотные остатки, принимающие участие в акте катализа.

29
Белковая часть сложных ферментов каталитически неактивна и называется апоферментом. Связывание апофермента с небелковым компонентом приводит к образованию каталитически активного фермента (холофермента):
:")
30
Некоторые ферменты наряду с активным центром содержат аллостерический (регуляторный) центр - участок белковой глобулы, вне активного центра, где могут связываться вещества, регулирующие ферментативную активность. Эти вещества называют аллостерическими эффекторами (аллостерическими активаторами или ингибиторами).
 центр - участок белковой глобулы, вне активного центра, где могут связываться вещества, регулирующие ферментативную активность. Эти вещества называют аллостерическим")
31
Специфичность действия. Специфичность связывания с субстратом определяется апоферментом. Специфичность действия ферментов определяет направленный обмен веществ в организме. О ферментах говорят, что они имеют узкую субстратную специфичность, если они действуют на очень небольшой круг субстратов. Иногда можно говорить об абсолютной субстратной специфичности, например, каталаза катализирует только одну реакцию - разложение пероксида водорода:

32
Для большинства ферментов характерна относительная (широкая, групповая) субстратная специфичность, когда они катализируют группу однотипных реакций. Например, алкогольдегидрогеназа катализирует превращения спиртов в альдегиды, причем в качестве субстратов могут выступать метанол, этанол, пропанол и другие спирты.
 субстратная специфичность, когда они катализируют группу однотипных реакций. Например, алкогольдегидрогеназа катализирует превращения спиртов в альдегиды, причем в качестве субст")
33
Ферменты наделены также стереохимической специфичностью: их активный центр распознает молекулы субстратов по пространственной конфигурации. Например, оксидазы L-аминокислот активны только в отношении L-аминокислот и совершенно не действуют на их D-аналоги. Фумараза катализирует превращение только фумаровой кислоты (транс-изомер), но не действует на малеиновую кислоту (цис-изомер):

34
Термолабильность. При повышении температуры на 10 о С скорость реакции увеличивается в 2-4 раза согласно правилу Вант-Гоффа. Однако при температурах выше 40 о С существенной становится денатурация ферментов, что приводит к уменьшению суммарной активности:

35
Зависимость скорости реакции от рН. Зависимость скорости ферментативной реакции от рН имеет колоколообразный вид. Значения рН, при которых наблюдается наиболее высокая скорость ферментативной реакции, называют оптимальными (рН-оптимум). Характер кривых и значение рН-оптимума зависит от природы заряженных групп субстрата и заряженных групп фермента (особенно тех, которые входят в активный центр). Оптимум рН для большинства ферментов лежит в пределах от 6,0 до 8,0
. Характер крив")
36
Графически зависимость скорости ферментативной реакции от концентрации субстрата представляет полуветвь гиперболы: Константа Михаэлиса (Km) - это концентрация субстрата, при которой скорость ферментативной реакции равна половине максимальной скорости (Vмах)
 - это концентрация субстрата, при которой скорость ферментативной реакции равна половине максимальной скорости")
37
График зависимости скорости реакции от концентрации субстрата, представленный в двойных обратных координатах:

Еще похожие презентации в нашем архиве:

 - это высокоспецифичные белки, выполняющие функции биологических катализаторов. Катализатор - это вещество, которое ускоряет.")

 36000, гемоглобина.")


")



;")





, состоящие из остатков аминокислот, которые соединены пептидной связью. Белки Протеины.")

 Совокупность протекающих в клетке химических превращений, обеспечивающих её рост,")


