Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Конформационные (пространственные) матрицы, или феномен белковой наследственности
 матрицы, или феномен белковой наследственности")
2
Центральная догма молекулярной биологии по Crick, 1958 с дополнением (Бондарев и др., неопубл.)
")
3
Полимеризационная модель образования амилоида Белок в нативной конформации Белок в прионной конформации («затравка») Белок в прионной конформации Белки в нативной конформации Белок в нативной конформации фибрилла а б по Shkundina et al., 2007
 Белок в прионной конформации Белки в нативной конформации Белок в нативной конформации фибрилла а б по Shkundina et al., 2007")
4
Неинфекционные амилоиды: -болезнь Паркинсона, Альцгеймера, Хантингтона, диабет 2-го типа и т.д. - «функциональные амилоиды» (стресс-гранулы, долговременная память, противовирусный иммунный ответ и т.д.) Инфекционные амилоиды - прионы: -куру,болезнь Крейтцфельдта-Якоба (БКЯ),фатальная семейная бессонница, скрепи, или почесуха, овец, губчатая энцефалопатия крупного рогатого скота - прионы низших грибов
 Инфекционные амилоиды - прионы: -куру,болезнь К")
5
Неинфекционные амилоиды Белок α-синуклеин (ген SNCA), участвует в формировании синаптических везикул для секреции дофамина образует амилоиды – тельца Леви при болезни Паркинсона
, участвует в формировании синаптических везикул для секреции дофамина образует амилоиды – тельца Леви при болезни Паркинсона")
6
Неинфекционные амилоиды Пептид Aβ (ген APP, картирован в 21-й хромосоме) – функция неизвестна Белок tau (ген MAPT) – поддерживает стабильность цитоскелета нейронов Накопление агрегатов белка Aβ и tau характерно при болезни Альцгеймера
 – функция неизвестна Белок tau (ген MAPT) – поддерживает стабильность цитоскелета нейронов Накопление агрегатов белка Aβ и tau характерно при болезни Альцгеймера")
7
Неинфекционные амилоиды Пептид амилин (ген IAPP), секретируется вместе с инсулином, участвует в метаболизме сахаров Образование агрегатов амилина приводит к апоптозу β-клеток поджелудочной железы и развитию диабета 2-го типа Белок хантингтин (ген HTT), функция неизвестна, ассоциирован с микротрубочками и везикулами Накопление агрегатов хантингтина характерно для болезни Хантингтона
, секретируется вместе с инсулином, участвует в метаболизме сахаров Образование агрегатов амилина приводит к апоптозу β-клеток поджелудочной железы и развитию диабета 2-го типа Белок хантингтин (ген HTT")
8
Инфекционные амилоиды - прионы Прионы – инфекционные агенты, способные передаваться от больного организма здоровому Причина прионных заболеваний человека и животных – прионизация белка PrP (от Prion Protein). PrP – трансмембранный белок, поддерживает миелиновую оболочку аксонов Различные заболевания связаны с различными изоформами PrP Прионные заболевания -Куру - Болезнь Крейтцфельдта-Якоба -Фатальная семейная бессонница -Губчатая энцефалопатия крупного рогатого скота («бешенство коров») -Скрепи, или «почесуха», овец -Трансмиссивная энцефалопатия норок -Хроническая изнуряющая болезнь (олени и лоси)
. PrP – трансмембранный белок, поддерживает")
9
Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на средах, богатых соединениями азота URE2 (репрессор генов, контролирующих усвоение бедных источников азота, в присутствии богатых) Lacroute, 1971; Wickner, 1994 [PIN+] Иницация [PSI+] de novoRNQ1 (функция неизвестна)Derkatch et al., 2001 [ISP+] Подавление нонсенс- супрессии, вызванной мутациями SUP35 SFP1 (транскрипционный фактор)Rogoza et al., 2010 [SWI+] Угнетение роста на средах, не содержащих глюкозу в качестве источника углерода SWI1/SNF5 (ремоделирование хроматина) Du et al., 2008 [OCT+] Способность использовать лактат в качестве единственного источника углерода на фоне о мутаций в гене CYC1 CYC8 (SSN6) (транскрипционный фактор) Patel et al., 2009 [MOT3] Потеря функции белка Mot3MOT3 (транскрипционный фактор)Alberti et al., 2009 [MOD5] Устойчивость к антимикотическим препаратам MOD5 (тРНК-изопентенил-трасфераза, модификация оснований в тРНК) Suzuki et al., 2012 [NUP100+] Не обнаруженоNUP100 (Нуклеопорин)Halfmann et al., 2012 [Het-s] * Несовместимость при слиянии гиф HET-sCoustou et al., 1997
![Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда](http://images.myshared.ru/5/395201/slide_9.jpg "Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда")
10
Критерии прионного наследования 1. Обратимая излечиваемость Если под воздействием некоторых агентов возможно излечивание клеток от приона, то он может возникнуть вновь в «излеченном» штамме 2. Сверхпродукция нормального клеточного белка увеличивает частоту появления приона В результате сверхпродукции нормального клеточного белка появляется больше молекул, способных спонтанно перейти в прионную форму. Это увеличивает частоту спонтанного возникновения прионной формы 3. Для поддержания приона необходимо присутствие нормального клеточного белка. Wickner, 1996

11
Дрожжевой прион [PSI+] супрессия Функциональный белок Sup35 узнает стоп- кодон, возникший на месте значащего Синтезируется укороченный нефункциональный белок Ade1 и дрожжи не растут на среде без аденина Синтезируется полный функциональный белок Ade1 и дрожжи растут на среде без аденина Прионные агрегаты Sup35 не узнают стоп- кодон, и он прочитывается как значащий
![Дрожжевой прион [PSI+] супрессия Функциональный белок Sup35 узнает стоп- кодон, возникший на месте значащего Синтезируется укороченный нефункциональный белок Ade1 и дрожжи не растут на среде без аденина Синтезируется полный функциональный белок Ade1](http://images.myshared.ru/5/395201/slide_11.jpg "Дрожжевой прион [PSI+] супрессия Функциональный белок Sup35 узнает стоп- кодон, возникший на месте значащего Синтезируется укороченный нефункциональный белок Ade1 и дрожжи не растут на среде без аденина Синтезируется полный функциональный белок Ade1")
12
Liebman S W, Chernoff Y O Genetics 2012;191: Copyright © 2012 by the Genetics Society of America Дрожжевой прион [PSI+]
![Liebman S W, Chernoff Y O Genetics 2012;191:1041-1072 Copyright © 2012 by the Genetics Society of America Дрожжевой прион [PSI+]](http://images.myshared.ru/5/395201/slide_12.jpg "Liebman S W, Chernoff Y O Genetics 2012;191:1041-1072 Copyright © 2012 by the Genetics Society of America Дрожжевой прион [PSI+]")
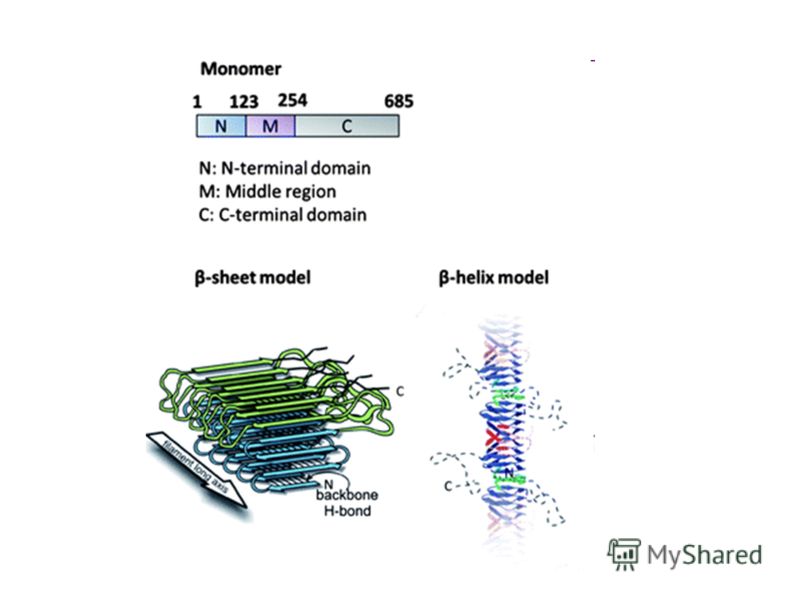
13
Структура белка Sup35

14
Дрожжевой прион [PSI+]
![Дрожжевой прион [PSI+]](http://images.myshared.ru/5/395201/slide_14.jpg "Дрожжевой прион [PSI+]")
15
Прион [PIN+]. Прионные сети [PIN+] – прион, необходимый для возникновения приона [PSI+] Агрегаты Rnq1 в клетках [PIN+] Прионные сети или каскад прионизации, - это отражение реальности системы протеома в клетке, когда изменения одного белка сказываются на структуре и функциях других белков. Следовательно, прионизация - это взаимодействие не только гомологичных, но и гетерологичных полипептидов. Прионизация Sup35 невозможна без взаимодействия этого белка с прионными агрегатами [PIN+] или [PSI+]. Поэтому в штаммах [psi-] прионные агрегаты [PIN+] играют роль затравки для прионизации Sup35
![Прион [PIN+]. Прионные сети [PIN+] – прион, необходимый для возникновения приона [PSI+] Агрегаты Rnq1 в клетках [PIN+] Прионные сети или каскад прионизации, - это отражение реальности системы протеома в клетке, когда изменения одного белка сказываютс](http://images.myshared.ru/5/395201/slide_15.jpg "Прион [PIN+]. Прионные сети [PIN+] – прион, необходимый для возникновения приона [PSI+] Агрегаты Rnq1 в клетках [PIN+] Прионные сети или каскад прионизации, - это отражение реальности системы протеома в клетке, когда изменения одного белка сказываютс")
16
Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на средах, богатых соединениями азота URE2 (репрессор генов, контролирующих усвоение бедных источников азота, в присутствии богатых) Lacroute, 1971; Wickner, 1994 [PIN+] Иницация [PSI+] de novoRNQ1 (функция неизвестна)Derkatch et al., 2001 [ISP+] Подавление нонсенс- супрессии, вызванной мутациями SUP35 SFP1 (транскрипционный фактор)Rogoza et al., 2010 [SWI+] Угнетение роста на средах, не содержащих глюкозу в качестве источника углерода SWI1/SNF5 (ремоделирование хроматина) Du et al., 2008 [OCT+] Способность использовать лактат в качестве единственного источника углерода на фоне о мутаций в гене CYC1 CYC8 (SSN6) (транскрипционный фактор) Patel et al., 2009 [MOT3] Потеря функции белка Mot3MOT3 (транскрипционный фактор)Alberti et al., 2009 [MOD5] Устойчивость к антимикотическим препаратам MOD5 (тРНК-изопентенил-трасфераза, модификация оснований в тРНК) Suzuki et al., 2012 [NUP100+] Не обнаруженоNUP100 (Нуклеопорин)Halfmann et al., 2012 [Het-s] * Несовместимость при слиянии гиф HET-sCoustou et al., 1997
![Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда](http://images.myshared.ru/5/395201/slide_16.jpg "Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда")
17
Прионы дрожжей и шапероны Hsp104 – основная дезагрегаза в клетках дрожжей, необходим для поддержания прионов. Делеция или сверхэкспрессия гена HSP104 приводит к потере [PSI+] Шапероны – белки, обеспечивающие правильную третичную укладку белков Hsp70 (Ssa и Ssb) – Ssa связываются с агрегатами в соотношении 1:2 и экранируют их от действия Hsp104 Ssb – стабилизируют неприонную форму мономеров Sup35 Кошапероны семейств Hsp40, Hsp90, Hsp110 – осуществляют тонкую настройку функций шаперонов семейства Hsp70, а также являются амилоидными «сенсорами» Модель организации агрегатов белка Sup35 в [PSI+] клетках (по Bagriantsev et al., 2008).
![Прионы дрожжей и шапероны Hsp104 – основная дезагрегаза в клетках дрожжей, необходим для поддержания прионов. Делеция или сверхэкспрессия гена HSP104 приводит к потере [PSI+] Шапероны – белки, обеспечивающие правильную третичную укладку белков Hsp70](http://images.myshared.ru/5/395201/slide_17.jpg "Прионы дрожжей и шапероны Hsp104 – основная дезагрегаза в клетках дрожжей, необходим для поддержания прионов. Делеция или сверхэкспрессия гена HSP104 приводит к потере [PSI+] Шапероны – белки, обеспечивающие правильную третичную укладку белков Hsp70")
18
Агрегаты α-синуклеина и шапероны Шапероны семейства Hsp 70 (Hsc70) и Hsp40 (Hdj) обеспечивают медленную разборку агрегатов α-синуклеина HspB5 готовит фибриллы α-синуклеина к разборке остальными шаперонами Hsp110 (Apg-2) и шаперон HspB5 усиливает действие шаперонов семейств Hsp70 и Hsp40 Добавление in vitro в эту систему Hsp104, значительно интенсифицирует процесс деполимеризации фибрилл
 и Hsp40 (Hdj) обеспечивают медленную разборку агрегатов α-синуклеина HspB5 готовит фибриллы α-синуклеина к разборке остальными шаперонами Hsp110 (Apg-2) и шаперон HspB5 усиливает дейс")
19
Поиск новых прионов in silico Michelitsch and Weissman, 2000 Алгоритм DIANA (Defined Interval Amino acid Numerating Algoritm), осуществляет поиск белков из дрожжевых баз данных, содержащих последовательности, обогащенные аспарагином и глутамином (108 белков, в том числе Swi1, Cyc8)
, осуществляет поиск белков из дрожжевых баз данных, содержащих последовательности, обогащенные аспарагином и глутамином (108")
20
Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на средах, богатых соединениями азота URE2 (репрессор генов, контролирующих усвоение бедных источников азота, в присутствии богатых) Lacroute, 1971; Wickner, 1994 [PIN+] Иницация [PSI+] de novoRNQ1 (функция неизвестна)Derkatch et al., 2001 [ISP+] Подавление нонсенс- супрессии, вызванной мутациями SUP35 SFP1 (транскрипционный фактор)Rogoza et al., 2010 [SWI+] Угнетение роста на средах, не содержащих глюкозу в качестве источника углерода SWI1/SNF5 (ремоделирование хроматина) Du et al., 2008 [OCT+] Способность использовать лактат в качестве единственного источника углерода на фоне о мутаций в гене CYC1 CYC8 (SSN6) (транскрипционный фактор) Patel et al., 2009 [MOT3] Потеря функции белка Mot3MOT3 (транскрипционный фактор)Alberti et al., 2009 [MOD5] Устойчивость к антимикотическим препаратам MOD5 (тРНК-изопентенил-трасфераза, модификация оснований в тРНК) Suzuki et al., 2012 [NUP100+] Не обнаруженоNUP100 (Нуклеопорин)Halfmann et al., 2012 [Het-s] * Несовместимость при слиянии гиф HET-sCoustou et al., 1997
![Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда](http://images.myshared.ru/5/395201/slide_20.jpg "Прионы низших грибов – цитоплазматические детерминанты ПрионФенотипическое проявление Структурный ген (его функция)Источник [PSI+] Нонсенс-супрессияSUP35 (фактор терминации трансляции) Cox, 1965; Wickner, 1994 [URE3] Усвоение уреидосукцината на среда")
21
Поиск новых прионов in silico Harrison and Gerstein, 2003 Алгоритм LPS (Lowest-Probability Subsequences) оценивает сходство или отличие белков от прионогенных доменов известных прионов (Sup35, Ure2, Rnq1) (170 белков). Методика выделяет несколько мотивов: QN – области обогащенные глутамином и аспарагином DERK – области обогащенные заряженными аминокислотами VILM – гидрофобный мотив
 оценивает сходство или отличие белков от прионогенных доменов известных прионов (Sup35, Ure2, Rnq1) (170 белков). Методика выделяет несколько мот")
22
Поиск новых прионов in silico Alberti et al., 2009 Алгоритм основывается на поиске последовательностей, богатых аспарагином и глутамином, при этом длина таких последовательностей (ядер) должна быть не менее 60 аминокислот. Выявлено 200 потенциально прионогенных белков. Были получены химерные конструкции, содержащие прионные домены этих белков, сшитые с геном YFP и изучена способность этих конструкций к формированию агрегатов при сверхэкспрессии Получены конструкции состоящие из функционального демена белка Sup35С и прионные домены потенциально прионогенных белков и проверена способность таких конструкций к индукции [PSI+], подобно полноразмерной копии гена SUP35 Проверена способность белков-кандидатов образовывать прионы in vivo. Выявлен новый прион [MOT3+]
 должна быть не менее 60 аминокислот. Выявлено 200 потенциально п")
24
Пространственные матрицы: патология или новый тип регуляции? Прионы – молекулярная патология? - Амилоидозы млекопитающих - Снижение скорости роста в 54% случаев при появлении [PSI+] Адаптивная роль прионов? -Нонсенс-супрессия в клетках [PSI+] (кратковременное появление приона может приводить к прочтению нонсенс-мутаций, «запирающих» псевдогены, к прочтению естественных стоп-кодонов, нейтрализация негативных последствий мутаций) -N-домен Sup35 консервативен у разных организмов от дрожжей до млекопитающих -Обнаружение [PSI+] и [MOT3+] в природных изолятах -Swi1 является хроматин-ремодулирующим фактором, Sfp1, Mot3, Cyc8 – глобальные транскрипционные факторы регулирующие 5-10% дрожжевого генома -[MOD5] – прионизация белка Mot5 приводит к устойчивости к антимикотическим препаратам
![Пространственные матрицы: патология или новый тип регуляции? Прионы – молекулярная патология? - Амилоидозы млекопитающих - Снижение скорости роста в 54% случаев при появлении [PSI+] Адаптивная роль прионов? -Нонсенс-супрессия в клетках [PSI+] (кратко](http://images.myshared.ru/5/395201/slide_24.jpg "Пространственные матрицы: патология или новый тип регуляции? Прионы – молекулярная патология? - Амилоидозы млекопитающих - Снижение скорости роста в 54% случаев при появлении [PSI+] Адаптивная роль прионов? -Нонсенс-супрессия в клетках [PSI+] (кратко")
25
Стресс-гранулы в клетках млекопитающих Стресс-гранулы –рибонуклеопротеиновые комплексы, образующиеся в ответ на стресс для остановки трансляции (кроме мРНК шаперонов) Удаление прионизующего домена TIA-1 клетка теряет способность образовывать соответствующие комплексы при стрессе. При этом химерный белок TIA-1(PrdΔ)-Sup35N восстанавливает функцию TIA-1. Сигнал к образованию стресс-гранул – инактивация фактора инициации трансляции eIF2 (за счет фосфорилирования стресс-активируемыми киназами). К преинициирующему комплексу присоединяются TIA-1 и TIAR, которые связывают мРНК
 Удаление прионизующего домена TIA-1 клетка теряет способность образовывать соответс")
26
Долговременная память Материальный носитель памяти должен длительно сохранять определенные состояния, а молекулы РНК и белков - короткоживущие Носителем памяти может служить продукт прионного превращения клеточного белка, обеспечивающего синаптическую связь между нейронами (стабилен и постоянно образуется из молекул того же белка) Tompa et al., 1998 У Aplisia californica cохранение функционально активной последовательности взаимодействия определенных нейронов связано с активацией трансляции некоторых мРНК, накапливающихся в синаптических окончаниях. Важную роль играет белок CPEB – регулятор трансляции, стимулирующий полиаденилирование мРНК и активирующий их трансляцию (в том числе мРНК белков, обеспечивающих стабилизацию ионных каналов в синапсах) N-домен СРЕВ богат аспарагином и глутамином (48%) Агрегация СРЕВ активирует трансляцию «молчащей» мРНК

27
Долговременная память Aplysia californica

28
Drosophila melanogaster Долговременная память

29
Роль прионоподобных агрегатов в противовирусном иммунном ответе Белок MAVS локализован на поверхности митохондрий в клетках млекопитающих и способен к прионизации, что запускает реакцию врожденного иммунного ответа Данный механизм является ярким примером «прионных сетей»

30
Роль прионоподобных агрегатов в противовирусном иммунном ответе Onoguchi et al., 2010 При проникновении вируса в клетку молекулы геликазы RIG-1 (компонент противовирусной системы клетки) связываются с РНК вируса. Происходит убиквитинилирование CARD- доменов (caspase-recruitment domain) RIG-1. Этот комплекс является затравкой для MAVS, имеющего сходные домены. Агрегаты взаимодействуют с сигнальными белками цитоплазмы, запуская последующий каскад и активацию белков IRF (interferon regulatory factors) и NF-kB, в результате синтезируется интерферон β
 связываются с РНК вируса. Происходит убиквитинилирование CARD-")
31
Меланосомы Меланосомы – органеллы меланоцитов человека – пигментных клеток, в которых происходит образование меланина, содержат трансмембранный белок Pmel17, в результате протеолиза образуется пептид Мα, который связывает мономеры – предшественники меланина, выполняющие функцию затравки. Полимеризация меланина приводит к образованию фибрилл пигмента, сходных по свойствам с амилоидами – биополимера, защищающего клетку от ультрафиолета. Домен Pmel17, отвечающий за полимеризацию, богат пролином, серином и треонином

32
Спидроины– белки основной нити паутины

33
Амилоиды прокариот Амилоидные фибриллы, образующие матрикс вокруг клетки E.сoli – «curli» Образование биопленок, колонизация новых субстратов, клеточная адгезия Белок CsgA образуют фибриллы «кюрли», характеризуются высоким содержанием β- слоев (домен ответственный за образование фибрилл обогащен глицином, глутамином, аспарагином) У Salmonella spp. показано образование фибрилл «tafi», состоящих из мономера белка AgfA (гомолог CsgA) 1. Поверхность воздушных гиф и спор Sterptomyces coelicolor обладает гидрофобными свойствами за счет 8-и белков ChpA-ChpH - чаплинов. 2. Патогенные свойства Xanthomonas spp. (патоген табака, дестабилизирует клеточные мембраны) обусловлены белками – харпинами (в том числе HpaG) Еще два примера белков, обогащенных β-слоями, образующими фибриллы:

34
Распространение последовательностей, склонных к агрегации Амилоидизация – широко распространенное явление Исследования, проводимые in silico показывают, что практически все белки имеют одинаковое количество регионов, склонных к агрегации Все белки способны образовывать амилоиды в нефизиологических условиях Последовательности, способные к агрегации ограничены специальными блокаторами полимеризации (привратниками) – все заряженные аминокислоты. Такие аминокислоты распознаются шаперонами. «Привратники» - специальный механизм регуляции адаптивной белковой агрегации?

35
Структура амилоидов Общие свойства амилоидов: 1.Формирование протяженных неразветвленных фибрилл (диаметр 3-10 нм) 2.Амилоиды обогащены β-слоями 3.Устойчивы к воздействиям денатурирующих агентов и протеаз Модель суперскладчатой β-структуры (Kajava et al., 2004) – основная роль в стабилизации структуры вносят межмолекулярные взаимодействия β-слои отдельных мономеров ориентированы перпендикулярно оси фибриллы. Молекулы при этом располагаются «в регистре» (точно друг под другом) Стабилизация за счет водородных связей между гидрофорбными остатками аминокислот
 2.Амилоиды обогащены β-слоями 3.Устойчивы к воздействиям денатурирующих агентов и протеаз Модель суперскладчатой β-структуры (Kajava et")
36
Строение фрагмента β-амилоида. Структура состоит из слоев двух мономеров, между которыми формируется солевой мост (К28-D23)
")
Еще похожие презентации в нашем архиве:

 матрицы, или феномен белковой наследственности.")




 РНК-геномы кодируют несколько белков, среди которых РНК-зависимая РНК-полимераза (репликаза), способная синтезировать.")






 высокомолекулярные органические вещества, состоящие из соединённых в цепочку пептидной связью аминокислот. Википедия.")

 внутренняя среда живой или умершей клетки, кроме ядра и вакуоли, ограниченная плазматической.")



