Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите



1
Молекулярная биология для биоинформатиков Академический университет Ефимова Ольга Алексеевна

2
Эпигенетика «Генетика предполагает, а эпигенетика располагает». P. Medawar & J. Medawar Лекция 10

3
Центральная догма молекулярной биологии: ДНК РНК БЕЛОК Генотип фенотип ДНК ответственна за хранение, передачу и реализацию наследственной информации

4
4 Доимплантационное развитие человека День 2. Эмбрион в стадии дробления 4 бластомера День 3. Эмбрион на стадии дробления 8 клеток. День 4. Морула. День 5. Бластоциста День 1. Стадия зиготы

5
Классификация стволовых клеток человека в соответствии с потенциалом к дифференцировке (Filip et al., 2004) Типы стволовых клеток человека Способность к дифференцировке Стволовые клетки в организме человека Тотипотентные клетки Все эмбриональные и экстра- эмбриональные ткани Оплодотворённый ооцит Бластомеры 2 – 8 клеточной стадии. Плюрипотентные клеткиВсе типы клеток эмбриона Эмбриональные стволовые клетки Первичные половые клетки Клетки эмбриональных карцином Пролиферирующие клетки дифференцированных тканей взрослого организма Мульти потентные Способны дифференцироваться в нескольких направлениях. Гемопоэтические Мышечные Нервной ткани Кожи Эндотелия Кишечника Миокарда Мезенхимные стволовые клетки Уни потентные Способны дифференцироваться только в одном направлении. Волосяного фолликула Семенников Яичников
 Типы стволовых клеток человека Способность к дифференцировке Стволовые клетки в организме человека Тотипотентные клетки Все эмбриональные и эк")
6
Разные судьбы, функции, морфология, «способности» клеток при одинаковом генотипе

7
Предмет эпигенетики «Исследование причинных взаимодействий между генами и их продуктами, приводящих к формированию фенотипа» (Waddington, 1942). Конрад Уоддингтон ( ) Эпигенетика в дополнение к генетике: «исследует явления, при которых генетическая изменчивость не ведет к изменениям фенотипа, а фенотипическая изменчивость, в свою очередь, не всегда может быть объяснена нарушениями генотипа» (Jablonka, Lamb, 2002). Классическая генетика и генетика развития: Изучение связи между изменчивостью генотипа и фенотипа в онтогенезе. Генотип + эпигенотип = фенотип
. Конрад Уоддингтон (1905-1975) Эпигенетика в дополнение к генетике: «исследует явления, при которых генети")
8
Эпигенетическое наследование В более общем смысле, предметом эпигенетики являются явления, связанные с развитием различных фенотипов клеток или организмов на основе одного генотипа. В более узком смысле эпигенетика – раздел генетики, который изучает наследуемые изменения активности генов во время развития организма или деления клеток. Эпигенетическое наследование – наследование паттерна экспрессии генов.

9
Эпигенетическая регуляция - наследственные и ненаследственные изменения в экспрессии конкретного гена без каких-либо соответствующих структурных изменений в его нуклеотидной последовательности. Эпигенетические явления: импринтинг, эффект положения, особенности структурно-функциональной организации хроматина определенных хромосомных локусов, влияющие на экспрессию генов, интерференция РНК.

10
ДВА ВИДА ИНФОРМАЦИИ В ГЕНОМЕ Генетическая – закодированная в ДНК программа создания живого организма Эпигенетическая (динамическая) – как, где и когда должна быть реализована генетическая информация. Каждый вид информации обеспечен своими системами: Кодирования Хранения Передачи
 – как, где и когда должна быть реализована генетическая информация. Каждый вид информации обеспечен своими системами:")
11
Изменения Необратимы (мутации) Изменения первичной структуры ДНК Стабильно наследуемые Обратимы Не затрагивают изменений первичной структуры ДНК Бывают долговременные и кратковременные генетическиеэпигенетические
 Изменения первичной структуры ДНК Стабильно наследуемые Обратимы Не затрагивают изменений первичной структуры ДНК Бывают долговременные и кратковременные генетическиеэпигенетические")
12
Метилирование ДНК Модификации гистонов Молекулярные основы эпигенетики Эпигеном - это совокупность всех эпигенетических маркеров, обусловливающих паттерн экспрессию генов в данной клетке.

13
Посттрансляционные модификации гистонов

14
Гистоны Н2А, Н2В, Н3 и Н4 формируют октамерные структуры, вокруг которых закручивается нить ДНК, образуя таким образом нуклеосомы

15
Структура нуклеосомы Аминокислотые остатки гистонов могут подвергаться пост- трансляционным модификациям: ацетилированию, фосфорилированию, метилированию. Модификации аминокислотных остатков гистоновых белков происходят, в основном, в N- терминальных участках, которые расположены за пределами компактного октамера и подвергаются действию различных клеточных сигналов

16
В зависимости от типа и сайта модификаций аминокислотных остатков, каждая нуклеосома имеет свой «гистоновый код», регулирующий активность транскрипции

17
Ацетилирование и деацетилирование гистонов ацетилирование связано с активацией транскрипции белки, осуществляющие ацетилирование - гистоновые ацетилтрансферазы (НАТ); донор ацетильной группы – ацетил коА белки, осуществляющие деацетилирование – гистоновые деацетилазы (HDAC) Модель модификации гистонов: ДНК-связывающиеся активаторы привлекают НАТ для ацетилирования нуклеосомных гистонов, а репрессоры привлекают HDAC для деацетилирования гистонов. Эти события приводят к изменению структуры нуклеосом и активации или репрессии транскрипции соответственно.
; донор ацетильной группы – ацетил коА белки, осуществляющие деацетилирование – гисто")
18
Эффект ацетилирования – ослабление связи между ДНК и гистонами из-за изменения заряда, в результате чего хроматин становится доступным для факторов транскрипции Сайты ацетилирования: аминогруппы лизиновых остатков в составе боковой цепи гистона

19
Фосфорилирование и дефосфорилирование гистонов фосфорилирование связано с активацией транскрипции белки, осуществляющие фосфорилирование – протеинкиназами; донор фосфата – АТФ белки, осуществляющие дефосфорилирование – фосфатазы Сайты фосфорилирования: гидроксильные группы серина, треонина и тирозина. В результате фосфорилирования увеличивается негативный заряд.

20
Метилирование гистонов Метилируются -Лизин (моно-, ди- и триметилирование) -Агринин (моно- и диметилирование) Метилирование не приводит к изменению заряда модифицируемого остатка Эффекты метилирования в зависимости от сайта модификации и количества метильных групп: -Репрессия транскрипции -Активация транскрипции Регуляция транскрипции через молекулы-эффекторы
 -Агринин (моно- и диметилирование) Метилирование не приводит к изменению заряда модифицируемого остатка Эффекты метилирования в зависимости от сайта модификации и количества м")
21
Метилирование лизинов Осуществляют лизиновые метилтрансферазы - НКМТ SET-домен 6 наиболее хорошо описанных сайтов метилирования: на гистоне Н3 (К4, К9, К27, К36, К79) на гистоне Н4 (К20) Деметлирование лизинов LSD1 удаляет метильные группы с Н3К4 JHDM1 – H3K36me1 и me2, JHDM2A – H3K9m1 и me2, JHDM3A – H3K36me3, JMJD2A – H3K9me3. Донор метильной группы – S-аденозилметионин (SAM)
 на гистоне Н4 (К20) Деметлирование лизинов LSD1 удаляет метильные группы с Н3К4 JHD")
22
Модификации Роль в транскрипцииСайты модифицирования Группа 1 ацетилированиеактивацияН3 (К9, К14, К18, К56) Н4 (К5, К8, К12, К16) Н2А (?) Н2В (К6, К7, К16, К17) фосфорилированиеактивацияН3 (S10) метилированиеактивацияН3 (К4, К36, К79) репрессияН3 (К9, К27) Н4 (К20) Группа 2 убиквитинированиеактивацияН2В (К123) репрессияН2А (К119) сумоилированиерепрессияН3 (?) Н4 (К5, К8, К12, К 16) Н2А (К126) Н2В (К6, К7, К16, К17) Роль модификаций в регуляции транскрипции
 Н4 (К5, К8, К12, К16) Н2А (?) Н2В (К6, К7, К16, К17) фосфорилированиеактивацияН3 (S10) метилированиеактивацияН3 (К4, К36, К79) репрессияН3 (К9")
23
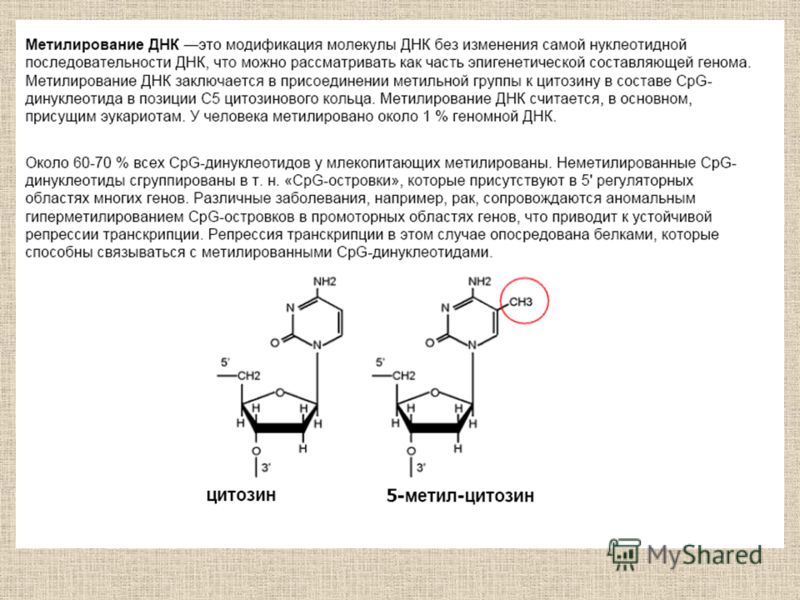
Метилирование ДНК и связанные с ним процессы

24
Молекулярные основы эпигенетики Б.Ф. Ванюшин Robin Holliday Впервые определил природу метилируемых последовательностей ДНК у разных видов организмов (1959 г.) Обосновал роль метилирования ДНК в регуляции работы гена. Предложил термин «эпимутация» (1987 г.) H N N N H O CH 3
 Обосновал роль метилирования ДНК в регуляции работы гена. Предложил термин «эпимутация» (19")
26
Репрессия транскрипции посредством метилирования ДНК

27
Взаимосвязь между метилированием цитозина в молекуле ДНК и ацетилированием гистонов

28
Механизмы инактивации гена в результате метилирования промоторной области 1. Метильные группы нарушают ДНК-белковые взаимодействия, выступая в большую бороздку ДНК и препятствуя связыванию специфических транскрипционных факторов. 2. Метилированные районы ДНК специфически связывают транскрипционные репрессоры. 3. Метилирование ДНК влияет на структуру хроматина.

29
Метилирование ДНК в клетке контролирует все (!) генетические процессы, в том числе такие как : Транскрипция (клеточная дифференцировка) Репликация Рекомбинация Репарация Транспозиция генов Инактивация Х-хромосомы
 генетические процессы, в том числе такие как : Транскрипция (клеточная дифференцировка) Репликация Рекомбинация Репарация Транспозиция генов Инактивация Х-хромосомы")
30
Резкое искажение метилирования ДНК: отсутствие метильных доноров (рак, гепатома) суперметилирование ДНК РАК полное выключение (knockout) ДНК-метилазного гена остановка развития, апоптоз, смерть (без метилирования ДНК жизни нет!) Биологическая специфичность метилирования ДНК: Видовая (штаммовая) Тканевая (клеточная) Органоидная (ядро, митохондрии, пластиды) Внутримолекулярная (островки метилирования, повторы) Возрастная
 суперметилирование ДНК РАК полное выключение (knockout) ДНК-метилазного гена остановка развития, апоптоз, смерть (без метилирования ДНК жизни нет!) Биологическая специфи")
31
Семейства ДНК-метилтрансфераз (ДНК-метилаз) млекопитающих: DNMT1 – поддержание метилирования В гаметогенезе изоформы: DNMT1o DNMTp DNMT2 – РНК-метилазная активность (может специфично метилировать цитозин в 38 положении антикодоновой петли тРНК аспарагина); связь между метаболическими процессами и репрограммированием метилирования ДНК DNMT3 – метилирование de novo, регуляторные функции при метилировании DNMT3a DNMT3b DNMT3L SAM – донор метильной группы
 млекопитающих: DNMT1 – поддержание метилирования В гаметогенезе изоформы: DNMT1o DNMTp DNMT2 – РНК-метилазная активность (может специфично метилировать цитозин в 38 положении антикодоновой петли тРНК аспара")
32
De novo метилирование ДНК и сохранение характера метилирования ДНК Высокометилированые последовательности: Сателлитная ДНК Повторяющиеся элементы (в т.ч. транспозоны и их инертные формы) Уникальная межгенная ДНК Экзоны генов
 Уникальная межгенная ДНК Экзоны генов")
33
Метилирование ДНК метафазных хромосом из мезенхимной стволовой клетки взрослого индивида лимфоцита плода человека 22/24 недель развития Клетки цитотрофобласта хориона легкого печени эмбриона человека 5/6 недель развития N=30 N=76 N=32 N=29 N=49

34
Локализация 5-метилцитозина на хромосоме 1. Оценка интенсивности флуоресценции гомологи хромосомы 1 АТ-5-МеСQFH G-сегментация АТ-5-МеС

35
CpG – островки -неметилированные участки длиной 1 kb - в 5`-концах 60% промоторов активных генов Что защищает их от метилирования? - они защищены белками - постоянная работа деметилаз - нетипичный состав оснований

36
Деметилирование – удаление метильных групп из ДНК Активное деметилирование – задействована ферментативная система, превращающая 5-метилцитозин в цитозин независимо от репликации Пассивное деметилирование – реализуется после репликации ДНК, за счет отсутствия метилазной активности. Новосинтезированная нить ДНК не метилируется по образцу старой, и образуется полуметилированная (гемиметилированная) ДНК. Долгое время механизм и ферменты, вовлеченные в процесс активного деметилирования ДНК оставались неизвестными!

37
Активное деметилирование ДНК 5-гидроксиметилцитозин – гидроксильная форма 5-метилцитозина может быть промежуточным соединением в процессе активного деметилирования (Tahiliani et al., 2009; Ito et al., 2010). 5-гидроксиметилцитозин описан у млекопитающих в начале 1970-х (Penn et al., 1972) год: 5-гидроксиметилцитозин выявлен в клетках: мозга почки легкого сердца в эмбриональных стволовых клетках мыши в клетках HeLa в клетках эмбриональной почки (Kriaucionis, Heintz, 2009; Tahiliani et al., 2009).
. 5-гидроксиметилцитозин описан у млекопитаю")
38
В 2009 году у млекопитающих было идентифицировано семейство белков TET (Ten-Eleven-Translocation), гомологичных белкам трипаносомы JBP1 и JBP2 – оксидазам метильной группы тимина (Tahiliani et al., 2009). Оказалось, что все три белка семейства TET – TET1, TET2 и TET3 – могут превращать 5-метилцитозин в 5-гидроксиметилцитозин (Ito et al., 2010). Активное деметилирование ДНК
, гомологичных белкам трипаносомы JBP1 и JBP2 – оксидазам метильной группы тимина (Tahiliani et al., 2009). Оказалось, что все три белка семейства TET –")
39
Деметилирование ДНК с образованием 5-гидроксиметилцитозина

40
Методы анализа метилирования 1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI) 2. Метилспецифическая ПЦР Трансформация цитозина в урацил бисульфитом Na 3. MethylLight – метилспецифическая ПЦР в реальном времени 4. Биологические микрочипы 5.Специфические антиметилцитозиновые антитела
 2. Метилспецифическая ПЦР Трансформация цитозина в урацил бисульфитом Na 3. MethylLight – метилспецифическая ПЦР в реальном времени 4. Биологические микрочипы 5.")
42
Волны эпигенетического репрограммирования генома млекопитающих ДНК примордиальных половых клеток значительно метилирована; при миграции клеток в недифференцированные гонады в них наблюдается резкое деметилирование; реметилирование (метилирование de novo) ДНК половых клеток происходит на поздних стадиях созревания. После оплодотворения уровень метилирования остается высоким в импринтированных генах, но резко снижается в неимпринтипрованных отцовских и материнских генах. К стадии бластоцисты уровень метилирования ДНК повышается.

43
Метаболизм SAM – донора метильной группы Метилирование ДНК и факторы внешней среды При дефиците фолиевой кислоты повышен риск возникновения дефектов нервной трубки у плода Причина: снижение уровня метилирования ДНК

44
Доказано влияние на метилирование ДНК металлов – никеля, кадмия, мышьяка, а также хрома, ртути, трихлорэтилена, дихлоруксусной и трихлоруксусной кислоты, бензола, бисфенола. Метилирование ДНК и факторы внешней среды Металлы способствуют образованию в клетке активных форм кислорода, вызывающих повреждения ДНК, которые затрудняют или делают невозможной работу ДНК метилтрансфераз. В 1992 году Баркером была выдвинута гипотеза FEBAD (fetal basis of adult disease). В пользу гипотезы свидетельствует обнаруженная взаимосвязь между воздействием на плод экзогенных и эндогенных факторов и риском последующего развития сердечно-сосудистых заболеваний, сахарного диабета второго типа, остеопороза и некоторых видов рака.

45
Внешние факторы, действующие на женщину в период беременности, могут изменять характер метилирования ДНК в ее клетках, модифицировать формирующиеся эпигенетические паттерны плода, а также влиять на процесс репрограммирования в формирующихся половых клетках плода! Внешние факторы

46
Метилирование ДНК является обратимой реакцией и в значительной степени подвержено воздействию эндогенных и экзогенных факторов. Эти особенности, с одной стороны, увеличивают риск возникновения ошибок из-за влияния негативных факторов, но с другой – дают возможность проводить коррекцию эпигенетической регуляции генома за счет определенных внешних воздействий, в том числе лекарственных средств, гормонов и диеты.

47
«В последние годы … установлен особый класс заболеваний человека, обусловленный дефектами структуры и модификаций хроматина - так называемые «хроматиновые болезни». С. Назаренко, 2005 г.

48
Синдром Ретта (OMIM ) dex.php?s=0b8265fee36f1322b6da b8dae8f038a7&showtopic=83503& pid= &st=765&#entry g/rettsyndrome.html 7_02_01_archive.html Частота 1 на детей женского пола Впервые описан Реттом в 1966г (Rett, 1966), повторно в 1983 Хогбергом ( Hagberg, 1993). Мутация в гене MeCP2 (MeC binding protein), расположенном на Xq28 регрессия развития аутизм стереотипные движения рук
 http://www.rodim.ru/conference/in dex.php?s=0b8265fee36f1322b6da b8dae8f038a7&showtopic=83503& pid=4926083&st=765&#entry49260 83 http://www.mississippichallenge.or g/rettsyndrome.html http://swimpig.blogspot.com/200 7_02_0")
49
Синдром ICF (OMIM ) ( Immunodeficiency, Centromere instability and Facial anomalies syndrome ) Мутации в гене DNMT3B (DNA metiltransferase), расположенном на хромосоме 20q11.2 Luciani et al., 2005 Впервые синдром описан в 1978 году (Hulten, 1978) Синдром ICF (иммунодефицит, хромосомная нестабильность, аномалии лицевого черепа) Гетерохроматиновые районы хромосом 1, 9 и 16 неметелированы, вследствие чего растянуты и имеют ветвистую структуру
 ( Immunodeficiency, Centromere instability and Facial anomalies syndrome ) Мутации в гене DNMT3B (DNA metiltransferase), расположенном на хромосоме 20q11.2 Luciani et al., 2005 Впервые синдром описан в 1978 году (Hulten, 197")
50
Синдром Коффина – Лоури (OMIM ) Мутация гена RSK (ribosomal S6 kinase), расположенном на Хp RSK2 - регулируемая ростовыми факторами серин- треониновая киназа book=gene&part=cls&rendertype=figure&id=cl s.F1 Частота встречаемости 1: Впервые был описан 1966 Коффином (Coffin et al., 1966), позже Лоури отметил другие характерные особенности в 1972 году (Lowry et al., 1972).
 Мутация гена RSK (ribosomal S6 kinase), расположенном на Хp21.1-21.2 RSK2 - регулируемая ростовыми факторами серин- треониновая киназа http://clsf.info/Welcome.htmhttp://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi? bo")
Еще похожие презентации в нашем архиве:

 Наследственные изменения в генах без изменения последовательности ДНК Сайленсинг генов.")


 Оперон (Ф.Жакоб, Ж.Л. Моно, 1961 г.) – группа генов, кодирующих белки,")







 Algorithms in Bioinformatics Analysis (www.saba.spbsu.ru) Синдром Ангельмана Курбацкий Евгений Николаев Павел Чернятчик.")





