Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Молекулярная биология для биоинформатиков Академический университет Ефимова Ольга Алексеевна

2
Эпигенетические модификации генома человека «Генетика предполагает, а эпигенетика располагает». P. Medawar & J. Medawar Лекция 2

3
3 Доимплантационное развитие человека День 2. Эмбрион в стадии дробления 4 бластомера День 3. Эмбрион на стадии дробления 8 клеток. День 4. Морула. День 5. Бластоциста День 1. Стадия зиготы

4
Разные судьбы, функции, морфология, «способности» клеток при одинаковом генотипе

5
Эпигенетическое наследование В более общем смысле, предметом эпигенетики являются явления, связанные с развитием различных фенотипов клеток или организмов на основе одного генотипа. В более узком смысле эпигенетика – раздел генетики, который изучает наследуемые изменения активности генов во время развития организма или деления клеток. Эпигенетическое наследование – наследование паттерна экспрессии генов.

6
Эпигенетическая регуляция - наследственные и ненаследственные изменения в экспрессии конкретного гена без каких-либо соответствующих структурных изменений в его нуклеотидной последовательности. Эпигенетические явления: импринтинг, эффект положения, особенности структурно-функциональной организации хроматина определенных хромосомных локусов, влияющие на экспрессию генов, интерференция РНК.

7
ДВА ВИДА ИНФОРМАЦИИ В ГЕНОМЕ Генетическая – закодированная в ДНК программа создания живого организма Эпигенетическая (динамическая) – как, где и когда должна быть реализована генетическая информация. Каждый вид информации обеспечен своими системами: Кодирования Хранения Передачи
 – как, где и когда должна быть реализована генетическая информация. Каждый вид информации обеспечен своими системами:")
8
Изменения Необратимы (мутации) Изменения первичной структуры ДНК Стабильно наследуемые Обратимы Не затрагивают изменений первичной структуры ДНК Бывают долговременные и кратковременные генетическиеэпигенетические
 Изменения первичной структуры ДНК Стабильно наследуемые Обратимы Не затрагивают изменений первичной структуры ДНК Бывают долговременные и кратковременные генетическиеэпигенетические")
9
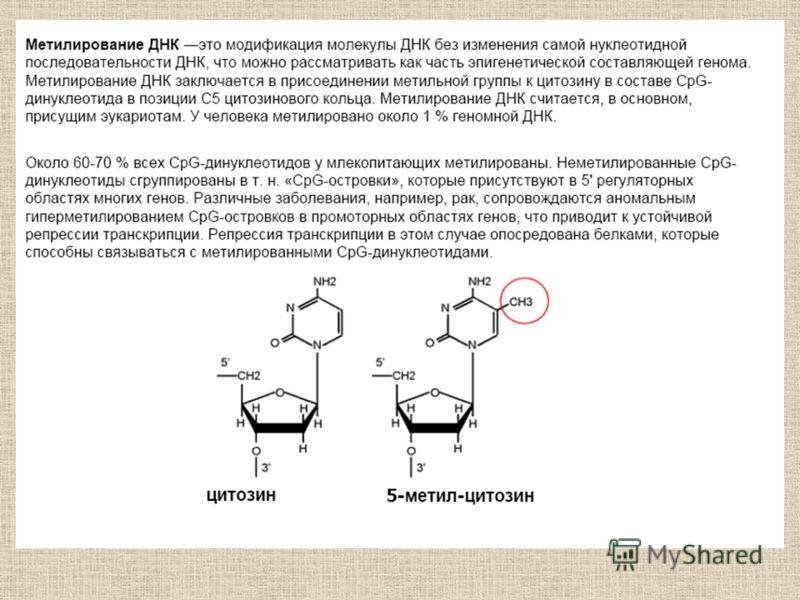
Метилирование ДНК Модификации гистонов Молекулярные основы эпигенетики Эпигеном - это совокупность всех эпигенетических маркеров, обусловливающих паттерн экспрессию генов в данной клетке.

10
Посттрансляционные модификации гистонов

11
Гистоны Н2А, Н2В, Н3 и Н4 формируют октамерные структуры, вокруг которых закручивается нить ДНК, образуя таким образом нуклеосомы

12
Структура нуклеосомы Аминокислотые остатки гистонов могут подвергаться пост- трансляционным модификациям: ацетилированию, фосфорилированию, метилированию. Модификации аминокислотных остатков гистоновых белков происходят, в основном, в N- терминальных участках, которые расположены за пределами компактного октамера и подвергаются действию различных клеточных сигналов

13
В зависимости от типа и сайта модификаций аминокислотных остатков, каждая нуклеосома имеет свой «гистоновый код», регулирующий активность транскрипции

14
Ацетилирование и деацетилирование гистонов ацетилирование связано с активацией транскрипции белки, осуществляющие ацетилирование - гистоновые ацетилтрансферазы (НАТ); донор ацетильной группы – ацетил коА белки, осуществляющие деацетилирование – гистоновые деацетилазы (HDAC) Модель модификации гистонов: ДНК-связывающиеся активаторы привлекают НАТ для ацетилирования нуклеосомных гистонов, а репрессоры привлекают HDAC для деацетилирования гистонов. Эти события приводят к изменению структуры нуклеосом и активации или репрессии транскрипции соответственно.
; донор ацетильной группы – ацетил коА белки, осуществляющие деацетилирование – гисто")
15
Эффект ацетилирования – ослабление связи между ДНК и гистонами из-за изменения заряда, в результате чего хроматин становится доступным для факторов транскрипции Сайты ацетилирования: аминогруппы лизиновых остатков в составе боковой цепи гистона

16
Фосфорилирование и дефосфорилирование гистонов фосфорилирование связано с активацией транскрипции белки, осуществляющие фосфорилирование – протеинкиназами; донор фосфата – АТФ белки, осуществляющие дефосфорилирование – фосфатазы Сайты фосфорилирования: гидроксильные группы серина, треонина и тирозина. В результате фосфорилирования увеличивается негативный заряд.

17
Метилирование гистонов Метилируются -Лизин (моно-, ди- и триметилирование) -Агринин (моно- и диметилирование) Метилирование не приводит к изменению заряда модифицируемого остатка Эффекты метилирования в зависимости от сайта модификации и количества метильных групп: -Репрессия транскрипции -Активация транскрипции Регуляция транскрипции через молекулы-эффекторы
 -Агринин (моно- и диметилирование) Метилирование не приводит к изменению заряда модифицируемого остатка Эффекты метилирования в зависимости от сайта модификации и количества м")
18
Метилирование лизинов Осуществляют лизиновые метилтрансферазы - НКМТ SET-домен 6 наиболее хорошо описанных сайтов метилирования: на гистоне Н3 (К4, К9, К27, К36, К79) на гистоне Н4 (К20) Деметлирование лизинов LSD1 удаляет метильные группы с Н3К4 JHDM1 – H3K36me1 и me2, JHDM2A – H3K9m1 и me2, JHDM3A – H3K36me3, JMJD2A – H3K9me3. Донор метильной группы – S-аденозилметионин (SAM)
 на гистоне Н4 (К20) Деметлирование лизинов LSD1 удаляет метильные группы с Н3К4 JHD")
19
Модификации Роль в транскрипцииСайты модифицирования Группа 1 ацетилированиеактивацияН3 (К9, К14, К18, К56) Н4 (К5, К8, К12, К16) Н2А (?) Н2В (К6, К7, К16, К17) фосфорилированиеактивацияН3 (S10) метилированиеактивацияН3 (К4, К36, К79) репрессияН3 (К9, К27) Н4 (К20) Группа 2 убиквитинированиеактивацияН2В (К123) репрессияН2А (К119) сумоилированиерепрессияН3 (?) Н4 (К5, К8, К12, К 16) Н2А (К126) Н2В (К6, К7, К16, К17) Роль модификаций в регуляции транскрипции
 Н4 (К5, К8, К12, К16) Н2А (?) Н2В (К6, К7, К16, К17) фосфорилированиеактивацияН3 (S10) метилированиеактивацияН3 (К4, К36, К79) репрессияН3 (К9")
20
Метилирование ДНК и связанные с ним процессы

21
Молекулярные основы эпигенетики Б.Ф. Ванюшин Robin Holliday Впервые определил природу метилируемых последовательностей ДНК у разных видов организмов (1959 г.) Обосновал роль метилирования ДНК в регуляции работы гена. Предложил термин «эпимутация» (1987 г.) H N N N H O CH 3
 Обосновал роль метилирования ДНК в регуляции работы гена. Предложил термин «эпимутация» (19")
23
Репрессия транскрипции посредством метилирования ДНК

24
Взаимосвязь между метилированием цитозина в молекуле ДНК и ацетилированием гистонов

25
Механизмы инактивации гена в результате метилирования промоторной области 1. Метильные группы нарушают ДНК-белковые взаимодействия, выступая в большую бороздку ДНК и препятствуя связыванию специфических транскрипционных факторов. 2. Метилированные районы ДНК специфически связывают транскрипционные репрессоры. 3. Метилирование ДНК влияет на структуру хроматина.

26
Метилирование ДНК в клетке контролирует все (!) генетические процессы, в том числе такие как : Транскрипция (клеточная дифференцировка) Репликация Рекомбинация Репарация Транспозиция генов Инактивация Х-хромосомы
 генетические процессы, в том числе такие как : Транскрипция (клеточная дифференцировка) Репликация Рекомбинация Репарация Транспозиция генов Инактивация Х-хромосомы")
27
Резкое искажение метилирования ДНК: отсутствие метильных доноров (рак, гепатома) суперметилирование ДНК РАК полное выключение (knockout) ДНК-метилазного гена остановка развития, апоптоз, смерть (без метилирования ДНК жизни нет!) Биологическая специфичность метилирования ДНК: Видовая (штаммовая) Тканевая (клеточная) Органоидная (ядро, митохондрии, пластиды) Внутримолекулярная (островки метилирования, повторы) Возрастная
 суперметилирование ДНК РАК полное выключение (knockout) ДНК-метилазного гена остановка развития, апоптоз, смерть (без метилирования ДНК жизни нет!) Биологическая специфи")
28
Семейства ДНК-метилтрансфераз (ДНК-метилаз) млекопитающих: DNMT1 – поддержание метилирования В гаметогенезе изоформы: DNMT1o DNMTp DNMT2 – РНК-метилазная активность (может специфично метилировать цитозин в 38 положении антикодоновой петли тРНК аспарагина); связь между метаболическими процессами и репрограммированием метилирования ДНК DNMT3 – метилирование de novo, регуляторные функции при метилировании DNMT3a DNMT3b DNMT3L SAM – донор метильной группы
 млекопитающих: DNMT1 – поддержание метилирования В гаметогенезе изоформы: DNMT1o DNMTp DNMT2 – РНК-метилазная активность (может специфично метилировать цитозин в 38 положении антикодоновой петли тРНК аспара")
29
De novo метилирование ДНК и сохранение характера метилирования ДНК Высокометилированые последовательности: Сателлитная ДНК Повторяющиеся элементы (в т.ч. транспозоны и их инертные формы) Уникальная межгенная ДНК Экзоны генов
 Уникальная межгенная ДНК Экзоны генов")
30
CpG – островки -неметилированные участки длиной 1 kb - в 5`-концах 60% промоторов активных генов Что защищает их от метилирования? - они защищены белками - постоянная работа деметилаз - нетипичный состав оснований

31
Деметилирование – удаление метильных групп из ДНК Активное деметилирование – задействована ферментативная система, превращающая 5-метилцитозин в цитозин независимо от репликации Пассивное деметилирование – реализуется после репликации ДНК, за счет отсутствия метилазной активности. Новосинтезированная нить ДНК не метилируется по образцу старой, и образуется полуметилированная (гемиметилированная) ДНК. Долгое время механизм и ферменты, вовлеченные в процесс активного деметилирования ДНК оставались неизвестными!

32
Активное деметилирование ДНК 5-гидроксиметилцитозин – гидроксильная форма 5-метилцитозина может быть промежуточным соединением в процессе активного деметилирования (Tahiliani et al., 2009; Ito et al., 2010). 5-гидроксиметилцитозин описан у млекопитающих в начале 1970-х (Penn et al., 1972) год: 5-гидроксиметилцитозин выявлен в клетках: мозга почки легкого сердца в эмбриональных стволовых клетках мыши в клетках HeLa в клетках эмбриональной почки (Kriaucionis, Heintz, 2009; Tahiliani et al., 2009).
. 5-гидроксиметилцитозин описан у млекопитаю")
33
В 2009 году у млекопитающих было идентифицировано семейство белков TET (Ten-Eleven-Translocation), гомологичных белкам трипаносомы JBP1 и JBP2 – оксидазам метильной группы тимина (Tahiliani et al., 2009). Оказалось, что все три белка семейства TET – TET1, TET2 и TET3 – могут превращать 5-метилцитозин в 5-гидроксиметилцитозин (Ito et al., 2010). Активное деметилирование ДНК
, гомологичных белкам трипаносомы JBP1 и JBP2 – оксидазам метильной группы тимина (Tahiliani et al., 2009). Оказалось, что все три белка семейства TET –")
34
Деметилирование ДНК с образованием 5-гидроксиметилцитозина

35
Методы анализа метилирования 1. Метилчувствительная ПЦР (Not1, Eag1, SacII, HpaII, HhaI) 2. Метилспецифическая ПЦР Трансформация цитозина в урацил бисульфитом Na 3. MethylLight – метилспецифическая ПЦР в реальном времени 4. Биологические микрочипы 5.Специфические антиметилцитозиновые антитела
 2. Метилспецифическая ПЦР Трансформация цитозина в урацил бисульфитом Na 3. MethylLight – метилспецифическая ПЦР в реальном времени 4. Биологические микрочипы 5.")
37
Волны эпигенетического репрограммирования генома млекопитающих ДНК примордиальных половых клеток значительно метилирована; при миграции клеток в недифференцированные гонады в них наблюдается резкое деметилирование; реметилирование (метилирование de novo) ДНК половых клеток происходит на поздних стадиях созревания. После оплодотворения уровень метилирования остается высоким в импринтированных генах, но резко снижается в неимпринтипрованных отцовских и материнских генах. К стадии бластоцисты уровень метилирования ДНК повышается.

38
Метаболизм SAM – донора метильной группы Метилирование ДНК и факторы внешней среды При дефиците фолиевой кислоты повышен риск возникновения дефектов нервной трубки у плода Причина: снижение уровня метилирования ДНК

39
Доказано влияние на метилирование ДНК металлов – никеля, кадмия, мышьяка, а также хрома, ртути, трихлорэтилена, дихлоруксусной и трихлоруксусной кислоты, бензола, бисфенола. Метилирование ДНК и факторы внешней среды Металлы способствуют образованию в клетке активных форм кислорода, вызывающих повреждения ДНК, которые затрудняют или делают невозможной работу ДНК метилтрансфераз. В 1992 году Баркером была выдвинута гипотеза FEBAD (fetal basis of adult disease). В пользу гипотезы свидетельствует обнаруженная взаимосвязь между воздействием на плод экзогенных и эндогенных факторов и риском последующего развития сердечно-сосудистых заболеваний, сахарного диабета второго типа, остеопороза и некоторых видов рака.

40
Внешние факторы, действующие на женщину в период беременности, могут изменять характер метилирования ДНК в ее клетках, модифицировать формирующиеся эпигенетические паттерны плода, а также влиять на процесс репрограммирования в формирующихся половых клетках плода! Внешние факторы

41
Метилирование ДНК является обратимой реакцией и в значительной степени подвержено воздействию эндогенных и экзогенных факторов. Эти особенности, с одной стороны, увеличивают риск возникновения ошибок из-за влияния негативных факторов, но с другой – дают возможность проводить коррекцию эпигенетической регуляции генома за счет определенных внешних воздействий, в том числе лекарственных средств, гормонов и диеты.

42
Интерференция РНК и регуляция экспрессии генов

43
Andrew Z. FireCraig C. Mello Нобелевская премия по физиологии и медицине 2006 год "RNA interference – gene silencing by double-stranded RNA"

44
Предположение Fire & Mello: Двуцепочечные РНК (dsRNA) – запускающий механизм (триггер) системы интерференции РНК.
 – запускающий механизм (триггер) системы интерференции РНК.")
45
Интерференция РНК – это замолкание (сайленсинг) генов, обеспечиваемое двуцепочечными молекулами РНК (dsРНК). Явление интерференции РНК открыто в 1998 г. у Nematoda (Fire et al., 1998)
 генов, обеспечиваемое двуцепочечными молекулами РНК (dsРНК). Явление интерференции РНК открыто в 1998 г. у Nematoda (Fire et al., 1998)")
46
dsРНК в раз более эффективно активируют систему интерференции РНК, по сравнению с ssРНК. Инъекция в C.elegans РНК мышечного белка

47
Fire A., Xu S.Q., Montgomery M.K., Kostas S.A., Driver S.E., Mello C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806–811 (1998). Эффект генетического сайленсинга посредством интерференции РНК: распространяется в тканях организма (!) наследуется (!!!)
. Эффект генетического сайленсинга посредством интерференции")
48
Механизм интерференции РНК при исследовании интерференции РНК у растений обнаружены короткие молекулы РНК, маркирующие ген, подверженный сайленсингу; в условиях in vitro воспроизведены биохимические реакции интерференции РНК. Малые интерферирующие РНК (siRNAs – short interfering RNAs) – класс нуклеотидных двуцепочечных РНК, образующихся из более длинных двуцепочечных РНК.

49
Общая схема регуляции генетической экспрессии за счет интерференции РНК (по Novina, Sharp, 2004)
")
50
Функции siРНК 1.Сайленсинг мобильных генетических элементов; 2.Сайленсинг гетерохроматиновых повторов; 3.Сайленсинг генетического материала вирусного происхождения; 4.Ограничение степени экспрессии гена в определенных тканях.

51
При выделение фракций коротких РНК (19-25 нуклеотидов) из различных организмов обнаружен еще один класс малых РНК – микроРНК. МикроРНК (miRNAs - micro RNAs) – класс нуклеотидных одноцепочечных РНК, закодированных в уникальных генах геномов многоклеточных организмов.
 из различных организмов обнаружен еще один класс малых РНК – микроРНК. МикроРНК (miRNAs - micro RNAs) – класс 19-25 нуклеотидных одноцепочечных РНК, закодированных в уникальных генах геномов мног")
52
Схема образования miРНК (по Novina, Sharp, 2004)
")
53
Функция miРНК Обеспечивают сайленсинг различных генов, обычно, за счет частично комплементарного связывания с мРНК, в результате которого блокируется ее трансляция. один тип miРНК может регулировать трансляцию мРНК более 100 различных генов; степень ингибирования зависит от количества связывающихся miРНК (в 3UTR мРНК содержится несколько сайтов связывания).

54
Продукт dsРНК, закодированных в уникальных генах геномов многоклеточных организмов (>1% от всех генов у человека); мРНК может не разрушаться; Один тип miРНК регулирует разные гены. Продукт dsРНК, образующихся в результате транскрипции транспозонов, гетерохроматиновых повторов или генетического материала вирусного происхождения ; мРНК разрушается; Один тип siРНК обычно регулирует только один тип мРНК. miРНКsiРНК Отличия miРНК и siРНК
; мРНК может не разрушаться; Один тип miРНК регулирует разные гены. Продукт dsРНК, образующихся в результате транскрипции транспозонов, г")
55
созданы библиотеки коротких РНК и ДНК- векторов, кодирующих короткие РНК, мишенями которых является около 8000 генов генома человека; внедряется в практику терапевтическое применение синтетических коротких РНК для целенаправленного подавления генетической экспрессии при некоторых заболеваниях.

56
«В последние годы … установлен особый класс заболеваний человека, обусловленный дефектами структуры и модификаций хроматина - так называемые «хроматиновые болезни». С. Назаренко, 2005 г.

57
Синдром Ретта (OMIM ) dex.php?s=0b8265fee36f1322b6da b8dae8f038a7&showtopic=83503& pid= &st=765&#entry g/rettsyndrome.html 7_02_01_archive.html Частота 1 на детей женского пола Впервые описан Реттом в 1966г (Rett, 1966), повторно в 1983 Хогбергом ( Hagberg, 1993). Мутация в гене MeCP2 (MeC binding protein), расположенном на Xq28 регрессия развития аутизм стереотипные движения рук
 http://www.rodim.ru/conference/in dex.php?s=0b8265fee36f1322b6da b8dae8f038a7&showtopic=83503& pid=4926083&st=765&#entry49260 83 http://www.mississippichallenge.or g/rettsyndrome.html http://swimpig.blogspot.com/200 7_02_0")
58
Синдром ICF (OMIM ) ( Immunodeficiency, Centromere instability and Facial anomalies syndrome ) Мутации в гене DNMT3B (DNA metiltransferase), расположенном на хромосоме 20q11.2 Luciani et al., 2005 Впервые синдром описан в 1978 году (Hulten, 1978) Синдром ICF (иммунодефицит, хромосомная нестабильность, аномалии лицевого черепа) Гетерохроматиновые районы хромосом 1, 9 и 16 неметелированы, вследствие чего растянуты и имеют ветвистую структуру
 ( Immunodeficiency, Centromere instability and Facial anomalies syndrome ) Мутации в гене DNMT3B (DNA metiltransferase), расположенном на хромосоме 20q11.2 Luciani et al., 2005 Впервые синдром описан в 1978 году (Hulten, 197")
59
Синдром Коффина – Лоури (OMIM ) Мутация гена RSK (ribosomal S6 kinase), расположенном на Хp RSK2 - регулируемая ростовыми факторами серин- треониновая киназа book=gene&part=cls&rendertype=figure&id=cl s.F1 Частота встречаемости 1: Впервые был описан 1966 Коффином (Coffin et al., 1966), позже Лоури отметил другие характерные особенности в 1972 году (Lowry et al., 1972).
 Мутация гена RSK (ribosomal S6 kinase), расположенном на Хp21.1-21.2 RSK2 - регулируемая ростовыми факторами серин- треониновая киназа http://clsf.info/Welcome.htmhttp://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi? bo")
60
Газета "Российское здоровье и общество" «Капризная дама» ГЕНЕТИКА: «Тричетыре года прошло, как появились хроматиновые болезни. Одна из них синдром Халькинсона-Килборга. Со времен вьетнамской войны его связывали с дефолиантами, применяемыми американской армией, затем с другими экстремальными воздействиями экологического характера. Причиной оказалась аномалия прикрепления хроматина к генам».

Еще похожие презентации в нашем архиве:



 Наследственные изменения в генах без изменения последовательности ДНК Сайленсинг генов.")
 Оперон (Ф.Жакоб, Ж.Л. Моно, 1961 г.) – группа генов, кодирующих белки,")


. 2.Двуцепочечная РНК действует (вызывает.")




 РНК-геномы кодируют несколько белков, среди которых РНК-зависимая РНК-полимераза (репликаза), способная синтезировать.")






