Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
LEGACY UNIVERSITY, THE GAMBIA Knowledge and Research for Integrity ANA 212 LECTURES WEEK FIVE SUMMER SEMESTER 2019/2020

2
Objectives Comprehension of the germ layer contributions to the early gastrointestinal tract (GIT) Comprehension of the folding of the GIT Comprehension of three main GIT embryonic divisions Comprehension of associated organ development (liver, pancreas, spleen) Some appreciation of the mechanical changes (rotations) during GIT development Some appreciation of gastrointestinal abnormalities
 Comprehension of the folding of the GIT Comprehension of three main GIT embryonic divisions Comprehension of associated organ development (liver, pancr")
3
The gastrointestinal tract (GIT) arises initially during the process of gastrulation from the endoderm of the trilaminar embryo (week 3) and extends from the buccopharyngeal membrane to the cloacal membrane. The tract and associated organs later have contributions from all the germ cell layers. During the 4th week three distinct regions (fore-, mid- and hind-gut) extend the length of the embryo and will contribute different components of the GIT. The large mid-gut is generated by lateral embryonic folding which "pinches off" a pocket of the yolk sac, the 2 compartments continue to communicate through the vitelline duct. The oral cavity (mouth) is formed following breakdown of the buccopharyngeal membrane (oropharyngeal or oral membrane) and contributed to mainly by the pharynx lying within the pharyngeal arches. Loss of buccopharyngeal membrane opens the tract to amniotic fluid through the remainder of development, and during the fetal period the amniotic fluid is actively swallowed. From the oral cavity the next portion of the foregut is initially the pharynx, a single gastrointestinal (oesophagus) and respiratory (trachea) common tube, that lies behind the heart. Note that the respiratory tract will form from a ventral bud arising at this level. This current page provides an introductory overview, use the links below for descriptions of specific components and regions as well as developmental abnormalities.
 arises initially during the process of gastrulation from the endoderm of the trilaminar embryo (week 3) and extends from the buccopharyngeal membrane to the cloacal membrane. The tract and associated organs later have")
6
Germ Layer Contributions Endoderm - Epithelium & Associated Glands Mesoderm (Splanchnic) - Mesentery, Connective Tissues, smooth Muscle, Blood Vessels Ectoderm (Neural Crest) - Enteric Nervous System (Neural Tube) - Extrinsic Innervation Both Endoderm & Mesoderm will contribute to Associated Organs. Note that in the historic texts the term entoderm is used to describe endoderm and other terminology may also differ from current descriptions.
 - Mesentery, Connective Tissues, smooth Muscle, Blood Vessels Ectoderm (Neural Crest) - Enteric Nervous System (Neural Tube) - Extrinsic Innervation Both Endoder")
8
Gastrointestinal Tract Divisions During the 4th week the 3 distinct divisions (foregut, Midgut and hindgut) extend the length of the embryo from oral membrane to cloacal membrane and will contribute different components of the GIT. These 3 divisions are also later anatomically defined by the vascular (artery) supply to each of theses divisions. The large mid-gut is generated by lateral embryonic folding which "pinches off" a pocket of the yolk sac, the 2 compartments continue to communicate through the vitelline duct. The oral cavity (mouth) is formed following breakdown of the buccopharyngeal membrane (oropharyngeal, oral membrane) and contributed to mainly by the pharynx lying within the pharyngeal arches. The opening of the GIT means that it contains amniotic fluid, which is also swallowed later in development.
 extend the length of the embryo from oral membrane to cloacal membrane and will contribute different components of the GIT. These 3 divisions")
10
First embryonic division of gastrointestinal tract, extending from the oral (buccopharyngeal) membrane and contributing oesophagus, stomach, duodenum (to bile duct opening), liver, biliary apparatus (hepatic ducts, gallbladder, and bile duct), and pancreas. The foregut blood supply is the celiac artery (trunk) excluding the pharynx, lower respiratory tract, and most of the oesophagus. From the oral cavity the next portion of the foregut is initially a single gastrointestinal (oesophagus) and respiratory (trachea) common tube, the pharynx which lies behind the heart. Note that the respiratory tract will form from a ventral bud arising at this level. Oral Cavity Pharynx (Esophagus, trachea) Respiratory Tract Stomach
 membrane and contributing oesophagus, stomach, duodenum (to bile duct opening), liver, biliary apparatus (hepatic ducts, gallbladder, and bile duct), and pa")
12
The middle embryonic division of gastrointestinal tract contributing the small intestine (including duodenum beneath distal bile duct opening), cecum, appendix, ascending colon, and part of the transverse colon (right half to two thirds). The midgut blood supply is the superior mesenteric artery. Much of the midgut is herniated at the umbilicus external to the abdomen through development. A key step in development is the rotation of this midgut that must occur to place the GIT in the correct abdominal position with its associated mesentry. The GIT itself differentiates to form significantly different structures along its length: oesophagus, stomach, duodenum, jejunum, ilium (small intestine), colon (large intestine). The mesenteries of the GIT are generated from the common dorsal mesentery, with the ventral mesentery contributing to the lesser omentum and falciform ligament. Note That the duodenum is commonly divided into 4 anatomical sequential parts (superior, descending, horizontal, ascending).
, cecum, appendix, ascending colon, and part of the transverse colon (right half to two thirds). The midgut")
21
Hindgut The final embryonic division of gastrointestinal tract consisting initially of the cloacae and extending to the cloacal membrane. The hindgut contributes part of the transverse colon (left half to one third), descending colon, sigmoid colon, rectum, part of anal canal (superior), urinary epithelium (bladder and most urethra). The initial cloaca space will later become partitioned by a septum into a dorsal gastrointestinal component (rectum) and ventral urogenital sinus (renal/genital component).
, descending colon, sigmoid colon,")
22
Carnegie stage 13 Embryo Overview OF THE GIT Below is an overview of the sections starting at the level of pharynx compressed dorsoventrally, following the GIT through to the rectum. The most obvious feature is that of a continuous tube initially, attached by dorsoventral mesentry. Outside this tube and mesentry (at the levels below the lung buds) is the intraembryonic coelom that will form the peritoneal cavity. The hepatic diverticulum (liver bud) lies under the septum transversum is the earliest associated GIT organ that has differentiated, and now occupies a substantial region of the abdomen.

23
Stomach curvature is generated by left-right asymmetric gut morphogenesis. Left-Right (LR) asymmetry is a fundamental feature of internal anatomy, yet the emergence of morphological asymmetry remains one of the least understood phases of organogenesis. Asymmetric rotation of the intestine is directed by forces outside the gut, but the morphogenetic events that generate anatomical asymmetry in other regions of the digestive tract remain unknown. Rodent experimental models have demonstrated that the mechanisms that drive the curvature of the stomach are intrinsic to the gut tube itself. The left wall of the primitive stomach expands more than the right wall, as the left epithelium becomes more polarized and undergoes radial rearrangement. These asymmetries exist across several species, and are dependent on LR patterning genes, including Foxj1, Nodal and Pitx2 Our findings have implications for how LR patterning manifests distinct types of morphological asymmetries in different contexts."
 asymmetry is a fundamental feature of internal anatomy, yet the emergence of morphological asymmetry remains one of the least understood phases of organogenesi")
24
Molecular The endoderm of the developing gastrointestinal tract is a source for patterning signals for both within the tract and also for the surrounding organs and tissues. Sox2 - Expressed in the anterior part of the primitive gut [ Cdx2 - Expressed in the posterior part of the primitive gut [7] [7] GDNF - Regulate migration of enteric neural crest cells [ Endothelin - Regulate migration of enteric neural crest cells [8] [8]

25
The Digestive Tract in Human Embryos Between Carnegie Stages 11 and 13 The digestive tract was initially formed by a narrowing of the yolk sac, and then several derived primordia such as the pharynx, lung, stomach, liver, and dorsal pancreas primordia differentiated during CS12 (21-29 somites) and CS13 ( 30 somites). The differentiation of four pairs of pharyngeal pouches was complete in all CS13 embryos. The respiratory primordium was recognized in 26-somite embryos and it flattened and then branched at CS13. The trachea formed and then elongated in 35-somite embryos. The stomach adopted a spindle shape in all 34-somite embryos, and the liver bud was recognized in 27-somite embryos. The dorsal pancreas appeared as definitive buddings in all but three CS13 embryos, and around these buddings, the small intestine bent in 33-somite embryos. In 35-somite embryos, the small intestine rotated around the cranial-caudal axis and had begun to form a primitive intestinal loop, which led to the Physiological umbilical herniation."

31
The gastrointestinal tract has both intrinsic and extrinsic innervation. The intrinsic innervation, the enteric plexus, is derived from neural crest cells migrating into and along the wall of the gastrointestinal tract. mainly vagal region neural crest - generating both neurons and glia. some sacral neural crest - in chicken. The extrinsic innervation occurs by efferent and afferent nerves, from the vagus and sympathetic chain and pelvic nerves. Vagus - Sensory and Motor Fibers project from oesophagus to small intestine. Sympathetic and Parasympathetic - lower oesophagus to large intestine. Pelvic Nerves - large intestine, rectum.

33
Gastrointestinal Tract Plexuses (Enteric Nervous System) Myenteric PlexusSubmucosal Plexus Auerbach's PlexusMeissner's Plexus Leopold Auerbach (1828–1897) A German Anatomist and Neuropathologist. Georg Meissner (1829–1905) a German Anatomist and Physiologist. First formed plexus Lies between the outer longitudinal and inner circular smooth muscle layers of muscularis externa Provides motor innervation to both layers Secretomotor innervation to the mucosa Both parasympathetic and sympathetic input Forms 2-3 days after the Myenteric Plexus Formed by Cells Migrating from the Myenteric Plexus Innervates Smooth Muscle of the Muscularis Mucosae Only Parasympathetic Fibers
 Myenteric PlexusSubmucosal Plexus Auerbach's PlexusMeissner's Plexus Leopold Auerbach (1828–1897) A German Anatomist and Neuropathologist. Georg Meissner (1829–1905) a German Anatomist and Phys")
34
Neural History 1857 Meissner was the first to describe a nerve plexus in the submucosa of the bowel wall Auerbach described the myenteric plexus between the longitudinal and circular muscle layers Le-Douarin describes neural crest contribution to both plexuses.

35
Myenteric Plexus Peristalsis Coordinated waves of descending inhibition followed by waves of descending excitation + Extrinsic parasympathetic cholinergic nerves (vagal and sacral) excite peristalsis and stimulate - Sympathetic noradrenergic nerves inhibit the transit of gut contents Submucosal Plexus Epithelial Movements Secretion and Absorption
 excite peristalsis and stimulate - Sympathetic noradrenergic nerves i")
36
Gastro-Intestinal Tract GIT Associated Organs Liver, pancreas and spleen (stage 22 embryo) See Legend Figure Immediately Below. The early tract develops as a simple tube, then a number of endodermal outgrowths from this tube at different levels and contribute to a range of additional organs and tissues. The gastrointestinal associated organs liver, gall bladder and pancreas. The Development of these organs is described on separate Slides below. There are also a number of additional non- gastrointestinal structures including the respiratory tract and development within the mesentery such as the spleen.
 See Legend Figure Immediately Below. The early tract develops as a simple tube, then a number of endodermal outgrowths from this tube at different levels and c")
38
Gastrointestinal Tract Abnormalities Only a brief description is given on this current page, for more details see Gastrointestinal Tract - Abnormalities. Lumen Abnormalities There are several types of abnormalities that impact upon the continuity of the gastrointestinal tract lumen. Atresia - interruption of the lumen (oesophageal atresia, duodenal atresia, extrahepatic biliary atresia, anorectal atresia) Stenosis - narrowing of the lumen (duodenal stenosis, pyloric stenosis). Duplication - incomplete recanalization resulting in parallel lumens, this is really a specialized form of stenosis. Meckel's Diverticulum This GIT abnormality is a very common and results from improper closure and absorption of the omphalomesenteric duct (vitelline duct) in development. This transient developmental duct connects the yolk to the primitive GIT.

40
Intestinal Aganglionosis (Intestinal Aganglionosis, Hirschsprung's disease, aganglionic colon, megacolon, congenital aganglionic megacolon, congenital megacolon) A condition caused by the lack of enteric nervous system (neural ganglia) in the intestinal tract responsible for gastric motility (peristalsis). Gastroschisis Gastroschisis (omphalocele, paraomphalocele, laparoschisis, abdominoschisis, abdominal hernia) is a congenital abdominal wall defect which results in herniation of fetal abdominal viscera (intestines and/or organs) into the amniotic cavity. Incidence of Gastroschisis has been reported at 1.66/10,000, occurring more frequently in young mothers (less than 20 years old). By definition, it is a body wall musculoskeletal defect, not a gastrointestinal tract defect, which in turn impacts upon GIT development.
 A condition caused by the lack of enteric nervous system (neural ganglia) in the intesti")
41
This section of the Lecture notes gives an overview of how the liver develops. Initially, the transverse septum (septum transversum) arises at an embryonic junctional site. The junctional region externally is where the ectoderm of the amnion meets the endoderm of the yolk sac. The junctional region internally is where the foregut meets the Midgut. The mesenchymal structure of the transverse septum provides a support within which both blood vessels and the liver begin to form. Arises at embryonic junction (septum transversum): externally is where ectoderm of amnion meets endoderm of yolk sac and internally is where the foregut meets the midgut. Mesenchymal structure of transverse septum provides a support within which both blood vessels and liver begin to form in the underlying splanchnic mesoderm. In the early embryo, the liver and heart grow rapidly forming obvious external swellings on the ventral embryo surface. The liver's initial embryonic function is mainly cardiovascular. Firstly, as a vascular connection between the developing placental vessels to the heart. Secondly, as a haemopoietic tissue where blood stem cells reside before bone marrow development. The fetal liver also has an endocrine role by 16-hydroxylation, that results in estriol being the major estrogen type produced in late human pregnancy. A recent molecular study has shown that within the adult liver at least 20 discrete cell populations exist these include: hepatocytes, endothelial cells, cholangiocytes, hepatic stellate cells, B cells, conventional and non-conventional T cells, NK-like cells, and distinct intrahepatic monocyte/macrophage populations.
 arises at an embryonic junctional site. The junctional region externally is where the ectoderm of the amnion meets th")
43
Human Embryonic Liver Development Week Carnegie Stage Feature Week 4 Carnegie stage 11 Hepatic diverticulum development (ductal plate) Carnegie stage 12 Cell differentiation Septum transversum forming liver stroma Hepatic diverticulum forming hepatic trabeculae Carnegie stage 13 Epithelial cord proliferation enmeshing stromal capillaries Week 5 Carnegie stage 14 Hepatic gland and its vascular channels enlarge Hematopoietic function appeared Week 7 Carnegie stage 18 Obturation due to epithelial proliferation Bile ducts became reorganized (continuity between liver cells and gut) Week 7 to 8 Carnegie stage to Carnegie stage 23 Biliary ductules developed in periportal Connective tissue Produces ductal plates that receive biliary capillaries
 Carnegie stage 12 Cell differentiation Septum transversum forming liver stroma Hepatic diverticulum forming hepatic")
44
Human data [11], see also liver development in the rat embryonic period (Carnegie stages 15-23). [12] (More? Detailed Timeline | Timeline human development) [11] [12]Detailed TimelineTimeline human development Embryonic Liver Development Timeline Carnegie Stage Age (days) CRL (mm) Biliary SystemVascularHepatic parenchyma Bile duct primordial duct links primitive intestine and liver parenchyma. Thick-walled tube (95 µm diameter) small lumen (22 µm diameter). Gall bladder - elongated tube further dilated, thick wall (125 µm diameter) and a narrow lumen (43 µm diameter). Hepatic sinusoids - intra-hepatic vasculature present Three venous tributaries flow into the liver sinusoids - right and left placental vein and a single vitelline vein. Cords of liver cells fragmented by vascular network of hepatic sinusoids. Between pericardial cavity (top) and mesonephros (bottom). Upper pole of the liver lies close to the septum transversum and early ventricles. Liver occupies the majority of abdominal cavity Bile duct (future common bile duct), and a common hepatic duct, in contact with liver parenchyma without penetration. Primordium of accessory bile tract is an elongated and fusiform gall bladder projecting forward and by a short cystic duct that opens into common bile duct. Bile duct empties into second part of duodenum on its posterior side. Portal system visible - portal vein (100 µm diameter) arises from connection of upper mesenteric vein then at region of hepatic hilum (285 µm) divides into portal branches. Left umbilical vein empties into anterior extremity of the left portal branch. Ductus venosus (80 µm) connects the initial portion of left portal vein to the inferior vena cava. Hepatic venous system 3 branches - left hepatic vein (120 µm in diameter), middle hepatic vein (220µm in diameter) and right hepatic vein (160 µm in diameter). Flows into the sub-cardinal vein. Liver parenchyma has two anatomical lobes (right and left lobe), separated by anteroposterior plane formed by placental vein Bile duct morphology as earlier stage. Common bile duct empties at the level of the proximal duodenum. Portal vein arises from the joining of splenic vein and superior mesenteric vein. At the level of the hepatic hilum, portal vein divides into two branches, right portal branch (420 µm in diameter) and left portal branch (540 µm in diameter). Right portal branch gives rise to a thin branch to caudate lobe. Ventral branch gives rise to segmental portal veins (VIII and V). Dorsal branch gives rise to the segmental portal veins (VI and VII). Ductus venosus connects initial portion of left portal vein to inferior vena cava, just upstream from hepatic vein afferents. Hepatic venous system as for previous stage. Hepatic parenchyma a large rounded mass Bile duct morphology as earlier stage. Portal venous system complete. Ductus venosus (40 µm) connects initial portion of portal vein to middle hepatic vein. Hepatic venous system has changed very little from the previous stage. Three hepatic veins empty into inferior vena cava. Liver parenchyma roughly oval shape, 2 symmetrical hepatic lobes. The quadrate and caudate lobes are identifiable. Upper pole of the liver bounded above by diaphragm.
![Human data [11], see also liver development in the rat embryonic period (Carnegie stages 15-23). [12] (More? Detailed Timeline | Timeline human development) [11] [12]Detailed TimelineTimeline human development Embryonic Liver Development Timeline Car](http://images.myshared.ru/118/1450027/slide_44.jpg "Human data [11], see also liver development in the rat embryonic period (Carnegie stages 15-23). [12] (More? Detailed Timeline | Timeline human development) [11] [12]Detailed TimelineTimeline human development Embryonic Liver Development Timeline Car")
45
Size - the liver initially occupies the entire anterior body area. Hepatoblast - endoderm the bipotential progenitor for both hepatocytes and cholangiocytes. Vascular - mesoderm blood vessels enter the liver (3 systems: systemic, placental, vitelline) Sinusoids - first blood vessels from vessels in septum transversum mesenchyme. Initially continuous endothelium, become fenestrated in fetal period and reticular development ongoing.

46
[1]. Hepatocytes - form 80% of liver, functional cells [2]. Cholangiocytes - epithelial cells that line the bile ducts [3]. Stellate Cells - mesenchymal cells in the space of Disse [4]. Kupffer Cells - liver macrophage in the sinusoids Summary Cell Map of the Adult Human Liver
![[1]. Hepatocytes - form 80% of liver, functional cells [2]. Cholangiocytes - epithelial cells that line the bile ducts [3]. Stellate Cells - mesenchymal cells in the space of Disse [4]. Kupffer Cells - liver macrophage in the sinusoids Summary Cell M](http://images.myshared.ru/118/1450027/slide_46.jpg "[1]. Hepatocytes - form 80% of liver, functional cells [2]. Cholangiocytes - epithelial cells that line the bile ducts [3]. Stellate Cells - mesenchymal cells in the space of Disse [4]. Kupffer Cells - liver macrophage in the sinusoids Summary Cell M")
48
Relevant Seminal studies has demonstrated that in the adult human liver at least twenty(20)discrete cell populations exist. These Include: Hepatocytes, Endothelial Cells, Cholangiocytes, Hepatic Stellate Cells, B-Cells, Conventional T-Cells and Non-Conventional T Cells, NK-Like Cells, and Distinct Intrahepatic Monocyte/Macrophage White Blood Cells Populations. Parenchymal Cells – such as Hepatocytes Non-Parenchymal Cells – such as Endothelial Cells, Cholangiocytes, Macrophages, Hepatic Stellate Cells, and Liver Infiltrating Lymphocytes-, Including B Cells, αβ and γδ, T Cells, and NK Cells. Non-inflammatory Macrophages are labeled Kupffer Cells based on their transcriptional similarity to mouse KC. The localizations of B Cells, Plasma Cells, T Cells, and NK Cells has yet to be confirmed by immunohistochemical staining of these populations in situ so their localization in the schematic Legend Figures beow is not representative of their zonated distribution.(Text Modified From Original Figure Legend)
discrete cell populations exist. These Include: Hepatocytes, Endothelial Cells, Cholangiocytes, Hepatic Stellate Cells, B-Cells, Conventional T-Cells and Non-C")
50
Liver Buds Differentiates to form the hepatic diverticulum and hepatic primordium, generates the gallbladder then divides into right and left hepatic (liver) buds. Three connecting stalks (cystic duct, hepatic ducts) which fuse to form bile duct. Left Hepatic Bud The Left Hepatic Bud Forms The : Left Lobe. The Left Lobe is made up of the: The Quadrate Lobe&The Caudate Lobe (both Q and C are Anatomically Left) The Caudate lobe of The Human Liver consists of Three(3) Anatomical parts: [1]-Spiegel's Lobe, [2]-Caudate Process& [3]-Paracaval Portion. Right Hepatic Bud The Right Hepatic Bud Forms The : Right Lobe
 buds. Three connecting stalks (cystic duct, hepatic ducts) which fuse to form bile duct. Lef")
52
Liver Structural Origins The Liver Lobule Hepatic Buds - Form hepatocytes, produce bile from week 13 (forms meconium of newborn) Vitelline Veins - Form sinusoids Mesenchyme - Form connective tissue and Kupffer cells Function - Haemopoiesis Embryonic liver also involved in blood formation, after the yolk sac and blood islands acting as a primary site.
 Vitelline Veins - Form sinusoids Mesenchyme - Form connective tissue and Kupffer cells Function - Haemopoiesis Embryonic")
54
Primitive Endoderm Foregut Diverticulum Foregut-Midgut Junction.Septum Transversum Hepatic Diverticulum Cystic Primordium GallBladder Common Bile Duct Hepatic Ducts Liver/GallBladder Hepatic Primordium Hepatic Parenchyma Hepatic Sinusoids Lobes of Liver Liver/GallBladder Midgut Region Hindgut Diverticulum (Pocket) Primary Data from Experimental Rodent Model Hepatoblasts - Endoderm-Derived Cells could Differentiate into either: 1.Hepatocytes - Populate the bulk of the Liver Parenchyma. 2.Cholangiocytes - Line the IntraHepatic Bile Ducts.

56
The transverse septum differentiates to form the hepatic diverticulum and the hepatic primordium, these two structures together will go on to form different components of the mature liver and gallbladder. At this stage large vascular channels can be seen coursing through the liver primordium.

58
The images below link to larger cross-sections of the mid-embryonic period (end week 4) stage 13 embryo starting just above the level of the liver and then in sequence through the liver to the level of the stomach. Note the relative position of the liver with respect to the abdominal cavity, the gallbladder and the heart.
 stage 13 embryo starting just above the level of the liver and then in sequence through the liver to the level of the stomach. Note the relative position of the l")
67
Cardiovascular ductus venosus dorsal aorta inferior vena cavaLiver Ductus Venosus Hepatocytes SinusoidsEndocrine Right Adrenal (Fetal Cortex) Left Adrenal (Fetal Cortex)Musculoskeletal Vertebral Body (Cartilage) Rib (Cartilage) Body Wall (Skeletal Muscle)Neural Spinal Cord Dorsal Root GangliaGastrointestinal Stomach (Pylorus)
 Left Adrenal (Fetal Cortex)Musculoskeletal Vertebral Body (Cartilage) Rib (Cartilage) Body Wall (Skeletal Mu")
68
The slides shown in the table below are to specific features shown on the Human embryo (stage 22) Liver and Ductus Venosus virtual slide. See also notes on Liver Development above & below
 Liver and Ductus Venosus virtual slide. See also notes on Liver Development above & below")
70
The images below depict larger cross-sections of the end of the embryonic period (week 8) stage 22 embryo starting just above the level of the liver and then in sequence through the entire liver. (Note the sections are viewed from below, LR axis is reversed) The rapidly developing liver also forms a visible surface bulge on the embryo directly under the heart bulge. The liver now occupies the entire ventral body cavity with parts of the gastrointestinal tract and urinary system embedded within its structure. Note in this image the large central ductus venosus.
 stage 22 embryo starting just above the level of the liver and then in sequence through the entire liver. (Note the sections are viewed from below, LR axis is r")
84
A - (Stain - Haematoxylin Eosin) Immuno-HistoChemistry B, C - Alpha-Fetoprotein (AFP) D, G - Cytokeratin 18 (CK18) E, H - Cytokeratin 19 (CK19) F, I - Cytokeratin 7 (CK7)
 Immuno-HistoChemistry B, C - Alpha-Fetoprotein (AFP) D, G - Cytokeratin 18 (CK18) E, H - Cytokeratin 19 (CK19) F, I - Cytokeratin 7 (CK7)")
86
Ductal Plate Development. The ductal plate is a primitive biliary epithelium which develops in mesenchyme adjacent to portal vein branches (Periportal Hepatoblasts). During liver development it is extensively reorganised (ductal plate remodelling) within the developing liver to form the intrahepatic bile ducts (IHBD). If remodelling does not occur, leading to excess of embryonic bile duct structures in the portal tract, these developmental abnormalities are described as ductal plate malformation (DPM). Cholangiocyte Tubulogenesis: Ductal Plate Stages -> Remodeling Bile Duct Stage -> Remodeled Bile Duct.
. During liver development it is extensively reorganised (ductal plate remodelling) wi")
88
Bile Secretion The epithelial cells that line the bile ducts are called cholangiocytes. The pathway below describes the production and passage of bile for final excretion into the duodenum: 1.hepatocytes produce bile 2.secreted into bile canaliculi 3.connected to intrahepatic bile ducts 4.intrahepatic bile ducts connect to the hepatic duct 5.then the cystic duct for storage in the gallbladder 6.then the common bile duct into the duodenum The term extrahepatic bile ducts (EHBDs) is used to describe the hepatic, cystic, and common bile ducts. The developing bile ducts express VEGF while hepatoblasts express angiopoietin-1, these two signals are thought to regulate arterial vasculogenesis and remodeling of the hepatic artery respectively.

89
Liver Vascular Development Data Venous week 4 - hepatic primordium in contact with vitelline veins and the umbilical veins. Week 4 to 6 - efferent venous vessels form from the vitelline veins. Afferent venous liver circulation present. Week 6 Onwards - Portal Vein formed from segments of the vitelline veins Portal Sinus (from Subhepatic Intervitelline Anastomosis) connects Umbilical vein to portal system. Ductus Venosus connects portal sinus to vena cava inferior. Birth - both umbilical vein and ductus venosus collapse. Portal vein becomes the only afferent vein of the liver. Arterial Week 8 - hepatic artery forms. Week 10 to 15 - intrahepatic arterial branches progressively extend from the central to the peripheral areas of the liver. Hepatic Sinusoids Week 4 - hepatic cords invade the septum transversum Progressively acquire structural and functional characters, through a multistage process.

91
Dual blood supply of the liver merges upon entry into the liver lobule at the portal field. The blood flows along the sinusoid and exits at the central vein. 1.branches of the portal vein 2.branches of the hepatic artery Portal Vein - is the sole supplier to the liver until about human 20 mm CRL stage. Portal vein primary branches extend around the periphery of each primitive liver lobule. This branching process continues, from primary to secondary, with each development supplying all newly forming liver lobules. Primary branches also lie parallel to the branches of hepatic veins, that drain the blood from the centre of each early lobule. Hepatic Artery - from the coeliac axis, initially contact the hepatic duct and gallbladder, later grows into the connective tissue about the larger bile ducts and branches of the portal vein. The hepatic artery will also supply the capsule of the liver.

92
These are the adult functional cells forming the majority of the liver (80% of the cells). Many Different Functions including: Storage of substances including glucose (as glycogen), vitamin A (possibly in specialized adipocytes), vitamin B12, folic acid and iron. Lipid Turnover synthesis of plasmalipoproteins Plasma Protein Synthesis albumin, alpha and beta globulins, prothrombin, fibrinogen Metabolism fat soluble compounds (drugs, insecticides), steroid hormones turnover Secretion bile (about 1 litre/day)
. Many Different Functions including: Storage of substances including glucose (as glycogen), vitamin A (possibly in specialized adipocytes), vitamin B12, folic a")
94
Kupffer Cells-Kupffer Cell Development Kupffer Cells are a population of tissue macrophages found in the lumen of hepatic sinusoids, their role is endocytic acting against blood-borne materials entering the liver. Primordial (primitive) macrophages arise in the yolk sac and then differentiate into fetal macrophages, either of these enter the blood and migrate into the developing liver. Tissue macrophages are a family of cells found in many organs: liver Kupffer cells, neural microglia, respiratory alveolar macrophages, and integumentary epidermal Langerhans cells. In the embryo, they have a common embryonic origin from yolk sac (YS) erythro-myeloid progenitors (EMPs). In the adult, they are self-maintained in tissues independently of hematopoietic stem cells (HSCs). Kupffer cell development
 macrophages a")
95
Ductus Venosus - shunts approximately half the umbilical vein blood flow directly to the inferior vena cava. Portal Vein - carries blood from the gastrointestinal tract and spleen to the liver. Left Umbilical Vein - carries oxygenated blood from the placenta to the embryo/fetus. Right Umbilical Vein - vessel degenerates leaving a single (left) umbilical vein. Vena Revehens - veins from the sinusoid vessels in the liver to the inferior vena cava, that later develop into the hepatic veins.

101
Abnormalities Structural developmental anomalies of gallbladder, bile ducts or liver Fibropolycystic Liver Disease Biliary atresia - Biliary atresia is a rare disease characterised by an inflammatory biliary obstruction of unknown origin that presents in the neonatal period. It is the most frequent surgical cause of cholestatic jaundice in this age group. Untreated, this condition leads to cirrhosis and death within the first years of life. Anomalous portal venous connection Congenital Absence of Portal Vein Congenital Absence of the Portal Vein (CAPV) is a rare abnormality where the intestinal and splenic venous drainage bypass the liver and drain directly into the systemic veins through various porto-systemic shunts. Links: Gastrointestinal Tract - Abnormalities Ductal Plate Malformations Interlobular bile ducts - autosomal recessive polycystic kidney disease Smaller interlobular ducts - von Meyenburg complexes Larger intrahepatic bile ducts - Caroli's disease Hepatobiliary cysts Fetal hepatic cysts are generally benign, with a low likelihood of associated anomalies of the hepatobiliary tract, abnormal liver function or clinical symptoms.[24]

102
Maternal Liver There are several maternal pregnancy-associated direct and indirect liver diseases, see recent review. Intrahepatic cholestasis of pregnancy (ICP) - most common liver disease in pregnancy, presenting mainly in the second and third trimesters, with pruritis, elevated serum bile acids, and abnormal liver function tests. Many Health Teams have developed guidelines to address the risks, diagnosis and management of ICP. Bile Acids levels greater than 10 μmol/L are a common diagnostic marker. Liver Function Tests - significant aminotransferase increase, serum bilirubin not usually raised. Hyperemesis Gravidarum The clinical term for severe form of nausea and vomiting, which are common symptoms of early pregnancy ( weeks). Causal Factors include increased human chorionic gonadotropin (hCG) and steroids, multiple pregnancy and vitamin deficiency. The condition can lead to dehydration, ketonuria, catabolism and may require hospitalisation. Acute Fatty Liver Disease Of Pregnancy (AFLP) Fatty infiltration of the liver, occurring usually in the third trimester though can also occur postnatally. Pre-Eclampsia HELLP - haemolysis (with a micoangiopathic blood smear), elevated liver enzymes (LFTs) and low platelet count.
 - most common liver disease in pregnancy, presenting mainly in the second and third trim")
103
Events In The Liver Development In Experimental Rodent Animal Models Stage One - liver bud undergoes growth and is the major site of haematopoiesis. Stage Two - bi-potential hepatoblasts differentiate into hepatocytes or biliary epithelial cells. Stage Three - ductal plate partially becomes bi-layered Stage Four - ductal plate remodelling, focal dilations appear between the two cell layers.

104
Adult Liver Transplants About 6,000 liver transplant operations are performed in the North America every year The main limitation on numbers are the availability of donor organs.

105
This section of notes gives an overview of gallbladder (gall bladder, gall-bladder) and billary tree development, histology and abnormalities associated with the biliary system. In the adult, the gallbladder is a site of bile salt storage and concentration, to then be released into the duodenum where they act to solubilize dietary lipids by their detergent effect. Bile salts are a cholesterol derivative (breakdown product). The transverse septum differentiates to form the hepatic diverticulum and the hepatic primordium, these two structures together will go on to form different components of the mature liver and gallbladder. The hepatic diverticulum divides into two parts: pars hepatica (larger cranial part, primordium of the liver) and pars cystica (smaller ventral invagination, primordium of gallbladder). The pars cystica vacuolates and expands, the stalk becoming the cystic duct. This structure is initially hollow, then solid (by proliferation of epithelial lining), and then recanalized occurs by vacuolation of this expanded epithelium. There are several opinions as to whether the duct has a solid phase or remains patent throughout development. Note that in some animals, for example horse and elephant, the gall bladder is normally absent.
 and billary tree development, histology and abnormalities associated with the biliary system. In the adult, the gallbladder is a site of bile salt storage and concent")
109
Gall-bladder Human Embryo (CRL) 7.5 mm - epithelium is surrounded by a layer of mesenchyma, and the entire structure is so imbedded in the under surface of the liver that it causes only a slight swelling of the peritoneal surface. Above and on the sides the mesenchyma is in direct relation with the hepatic trabecular, and it receives a few prolongations of the venous capillaries. Below it is covered by the peritoneal epithelium except on the left, where that layer is reflected to the abdominal walls in connection with the falciform ligament. In later stages the gall-bladder is separated from the hepatic trabecular on either side, and is attached to the liver only along its upper surface. 16 mm mesenchyma surrounding the gall-bladder is still undifferentiated mm forms two broad concentric zones, of which the inner is darker and more compact than the outer. 29 mm certain cells in the peripheral part of the dark zone form a third layer, which is thin and somewhat interrupted. As seen in later stages these cells are myoblasts, so that at 29 mm all three layers of the adult gall-bladder are indicated. These are the mucosa, muscularis, and serosa. The layers become gradually less distinct toward the hepatic duct.
 7.5 mm - epithelium is surrounded by a layer of mesenchyma, and the entire structure is so imbedded in the under surface of the liver that it causes only a slight swelling of the peritoneal surface. Above and on the si")
111
Studies of the Anatomy of rodent and human livers revealed some interesting differences. The size of the liver of terrestrial mammals obeys the allometric scaling law over a weight range of > Since scaling reflects adaptive changes in size or scale among otherwise similar animals, it could be expected to observe more similarities than differences between rodent and human livers. Obvious differences, such as the presence in (rodents) or the absence in (humans) of lobation and the presence in both (mice&humans) or the absence in (rats) of a gallbladder, suggest qualitative differences between the livers of these species. After review, however, it could be concluded that these dissimilarities represent relatively small quantitative differences. The microarchitecture of the liver is very similar among mammalian species and best represented by the lobular concept, with the biggest difference present in the degree of connective tissue development in the portal tracts. Although larger mammals have larger lobules, increasing size of the liver is mainly accomplished by increasing the number of lobules. The increasing role of the hepatic artery in lobular perfusion of larger species is, perhaps, the most important and least known difference between small and large livers, because it profoundly affects not only interventions like liver transplantations, but also calculations of liver function.

112
The fate of the vitelline and umbilical veins during the development of the human liver. Differentiation of endodermal cells into hepatoblasts is well studied, but the remodeling of the vitelline and umbilical veins during liver development is less well understood. Comparisons between human embryos between 3 and 10 weeks of development with pig and mouse embryos at comparable stages, and used Amira 3D reconstruction and Cinema 4D remodeling software for visualization. The vitelline and umbilical veins enter the systemic venous sinus on each side via a common entrance, the hepatocardiac channel. During expansion into the transverse septum at Carnegie Stage (CS)12 the liver bud develops as two dorsolateral lobes or wings and a single ventromedial lobe, with the liver hilum at the intersection of these lobes. The dorsolateral lobes each engulf a vitelline vein during CS13 and the ventromedial lobe both umbilical veins during CS14, but both venous systems remain temporarily identifiable inside the liver.

113
The dominance of the left-sided umbilical vein and the rightward repositioning of the sinuatrial junction cause de novo development of left-to-right shunts between the left umbilical vein in the liver hilum and the right hepatocardiac channel (venous duct) and the right vitelline vein (portal sinus), respectively. Once these shunts have formed, portal branches develop from the intrahepatic portions of the portal vein on the right side and the umbilical vein on the left side. The gall bladder is a reliable marker for this hepatic vascular midline. No evidence for large-scale fragmentation of embryonic veins as claimed by the vestigial theory was found. Instead and in agreement with the lineage theory, the vitelline and umbilical veins remained temporally identifiable inside the liver after being engulfed by hepatoblasts. In agreement with the hemodynamic theory, the left-right shunts develop de novo.

114
Fate mapping of gallbladder progenitors in posteroventral foregut endoderm of mouse early somite stage embryos. In early embryogenesis, the posteroventral foregut endoderm gives rise to the budding endodermal organs including the liver, ventral pancreas and gallbladder during early somitogenesis. Despite the detailed fate maps of the liver and pancreatic progenitors in the mouse foregut endoderm, the exact location of the gallbladder progenitors remains unclear. In this study, a DiI fate-mapping analysis using whole-embryo cultures of mouse early somite-stage embryos was performed. It was demonstrated that the majority of gallbladder progenitors in 9-11-somite-stage embryos are located in the lateral-most domain of the foregut endoderm at the first intersomite junction level along the anteroposterior axis. This definition of their location highlights a novel entry point to understanding of the molecular mechanisms of initial specification of the gallbladder.

115
Abnormalities Structural developmental anomalies of gallbladder Agenesis, aplasia or hypoplasia of gallbladder Bile Ducts Structural Developmental Anomalies Of Bile Ducts Choledochal Cyst Biliary Atresia Biliary atresia - Biliary atresia is a rare disease characterized by an inflammatory biliary obstruction of unknown origin that presents in the neonatal period. It is the most frequent surgical cause of cholestatic jaundice in this age group. Untreated, this condition leads to cirrhosis and death within the first years of life. Congenital stenosis or stricture of bile ducts (Congenital Hypoplasia Of Bile Ducts) Structural developmental anomalies of cystic duct Accessory bile duct Infection--Bacterial Infections These mainly relate to postnatal infections. Recent studies in the mouse have identified that gastrointestinal tract listeria infections can relocate to the gallbladder and reside there, leading to later reinfection of the gastrointestinal tract.

116
This section of the Lecture notes gives an overview of how the pancreas develops as an exocrine organ associated with the gastrointestinal tract. There is a second description, similar in overview, in relation to the pancreas as an endocrine organ, see Endocrine - Pancreas Development. At the foregut/midgut junction the septum transversum generates 2 pancreatic buds (dorsal and ventral endoderm) which will fuse to form the pancreas. The dorsal bud arises first and generates most of the pancreas. The ventral bud arises beside the bile duct and forms only part of the head and uncinate process of the pancreas. The digestive enzyme alpha amylase is the main exocrine enzyme produced by the pancreas (Amy2), and also by salivary glands (Amy1). Study of prenatal fetal and postnatal production of amylase during the first 2 years has shown that the majority of produced initially is salivary amylase. Infants and newborns also show a poor secretory response, to secretin, that is either absent or minimal at birth and is then acquired during the postnatal period. These results taken together suggest that the exocrine pancreas only becomes gradually functional postnatally.

119
Functions- exocrine (amylase, alpha-fetoprotein) and endocrine (pancreatic islets) Pancreatic buds- endoderm, covered in splanchnic mesoderm Ventral bud later (stage 13 - stage 14) Pancreatic bud formation - duodenal level endoderm, splanchnic mesoderm forms dorsal and ventral mesentery, dorsal bud (larger, first), ventral bud (smaller, later) Week 6 - Duodenum growth/rotation - brings ventral and dorsal buds together, fusion of buds Pancreatic Duct - ventral bud duct and distal part of dorsal bud, exocrine function Islet Cells- cords of endodermal cells form ducts, which cells bud off to form islets Week 7 to 20 - pancreatic hormones secretion increases, small amount maternal insulin Week 10 - glucagon (alpha) differentiate first, somatostatin (delta), insulin (beta) cells differentiate, insulin secretion begins
 and endocrine (pancreatic islets) Pancreatic buds- endoderm, covered in splanchnic mesoderm Ventral bud later (stage 13 - stage 14) Pancreatic bud formation - duodenal level endoderm, splanchnic mesode")
120
Week 15 - Glucagon detectable in Fetal Plasma Beta Cells – Stimulate Fetal Growth, continue to proliferate through to postnatal in infancy, most abundant. Maternal Diabetes Mellitus – Maternal Diabetes Mellitus leads to Hypertrophy Of Fetal Beta Cells.

121
During week 6 about day 41 (Carnegie stage 17, GA week 8) the stomach rotation brings the smaller ventral pancreatic bud dorsally to fuse with the larger dorsal pancreatic bud. Pancreatic Bud Rotation (Animal Models) Mouse - day 13 to 14 Pig - 38 to 52 days days post coitum (d.p.c.) Cattle - before days post coitum 45 (d.p.c.)
 the stomach rotation brings the smaller ventral pancreatic bud dorsally to fuse with the larger dorsal pancreatic bud. Pancreatic Bud Rotation (Animal Models) Mouse - day 13 to 14 Pig - 38 to")
124
Pancreatic Duct The initial formation of the pancreas as two separate lobes each with their own duct that fuses leads a range of anatomical variations in the adult exocrine pancreatic duct. The Five Variations In Pancreatic Duct Classification : common, ansa pancreatica, branch fusion, looped, and separated. Accessory pancreatic duct (APD, of Santorini) in the embryo is the main drainage duct of the dorsal pancreatic bud emptying into the minor duodenal papilla. In the adult it has been further classified as either long-type (joins main pancreatic duct at pancreas neck portion) and short-type (joins main pancreatic duct near first inferior branch).

125
Main Pancreatic Duct (MPD or Wirsung's duct) forms within the dorsal pancreatic bud and is present in the body and tail of the pancreas. Discovered by Johann Georg Wirsung ( ) a German physician who worked as a prosector in Padua. Accessory Pancreatic Duct (APD or Santorinis duct) is present mainly in the head of the pancreas. Originally dissected and delineated by Giovanni Domenico Santorini ( ) an Italian anatomist. Endoscopic Retrograde Cholangiopancreatography (ERCP) is a medical procedure which allows an injected dye to display the duct system on an x ray (pancreatograms).
 forms within the dorsal pancreatic bud and is present in the body and tail of the pancreas. Discovered by Johann Georg Wirsung (1589 - 1643) a German physician who worked as a prosector in Padua. Accessory")
128
Gastrointestinal Tract - Pancreas Histology Pancreas History main pancreatic duct (MPD) discovered by Johann Georg Wirsung ( ) a German physician who worked as a prosector in Padua. The duct is also called Wirsung's duct accessory pancreatic duct (APD) dissected and delineated by Giovanni Domenico Santorini ( ) an Italian anatomist. The duct is also called Santorini's duct Amylase, the form enzyme also found in exocrine pancreas, isolated from a malt solution by Anselme Payen Islets of Langerhans named in honour of Paul Langerhans ( ) by Gustave-Edouard Laguesse ( ) a french histopathologist discovery of insulin by Frederick Banting and John Macleod, two Canadian researchers, and they subsequently win the 1923 Nobel Prize in Medicine. Banting shared his part of the prize money with a younger coworker Charles Best glucagon, originally called "hyperglycemic glycogenolytic factor", purified by Staub, Sinn and Behrens.
 discovered by Johann Georg Wirsung (1589 - 1643) a German physician who worked as a prosector in Padua. The duct is also called Wirsung's duct. 1724 - acce")
131
Pancreas Digestive Pancreatic buds- endoderm, covered in splanchnic mesoderm Pancreatic bud formation - duodenal level endoderm, splanchnic mesoderm forms dorsal and ventral mesentery, dorsal bud (larger, first), ventral bud (smaller, later) Duodenum growth/rotation -ì brings ventral and dorsal buds together, fusion of buds Pancreatic duct - ventral bud duct and distal part of dorsal bud, exocrine function Functions - exocrine (amylase, alpha-fetoprotein) Pancreatic amylase digests starch to maltose. Postnatally, a blood test to detect amylase can be used to diagnose and monitor acute or chronic pancreatitis (pancreas inflammation). Pancreatic alpha-fetoprotein has been found to change in expression level (in rats) during developent and has been suggested to influence pancreas development. Immunolocalization for AFP revealed that a positive reactivity was detectable at E15.5 pancreas, became stronger in the cytoplasm of mesenchyme cells at E18.5, and declined after birth to a nearly undetectable level in adults." Secretagogues Secretagogue is a generic term for any substance that stimulates the secretion of another substance from a tissue or organ. In the exocrine pancreas, these are substances that stimulate the acini release of digestive enzymes. Note that bicarbonate and fluid secretion occurs in the pancreatic ducts and its stimulation may differ from enzyme secretion. secretin cholecystokinin vasoactive intestinal polypeptide (VIP)
, ventral bud (smaller, later) Duodenum")
132
After the stomach the initial portion of the gastrointestinal tract tube is the duodenum which initially lies in the midline within the peritoneal cavity. This region, along with the attached pancreas, undergoes rotation to become a retroperitoneal structure. This diagram shows the rotation with spinal cord at the top, vertebral body then dorsal aorta then pertioneal wall and cavity. Note this is a simplified diagram and the liver would push everything to the left during this rotation.

134
Experimental Rodent Pancreas Development Pancreas develops from distinct dorsal and ventral primordia. Dorsal pancreas - midline endoderm in posterior foregut is a single layer of epithelial cells that contacts notochord, an axial mesoderm-derived structure Ventral pancreas - Laterally, endoderm fated to form ventral pancreas is adjacent to both splanchnic mesoderm and aortic endothelial cells but is not in direct contact with notochord. The notochord and dorsal prepancreatic endoderms remain in contact until about the 13-somite stage in mice, 8.5 d postcoitum (dpc), when midline fusion of the paired dorsal aortas occurs. The first indication of morphogenesis occurs at somites in mice (9.5 dpc) Dorsal Mesenchyme condenses and underlying endoderm evaginates, forming a recognizable dorsal pancreatic bud The ventral bud appears later at ~30 somites ( dpc). Stimulated by mesenchymal signals, pancreatic epithelial cells proliferate and branch. The accumulated evidence is consistent with the possibility that a unique cell gives rise to all pancreatic cell lineages. The existence of such a pancreatic stem cell remains debatable.

136
Rodent Pancreas Cell Lineage In this study Rodent cell types were collected at different ages E11 and E15 pancreatic progenitors, E15 acinar cells, E15 endocrine progenitors (EP), E15, E17, P1, P15, 8–12 week beta cells, P1 and 8–12 week alpha cells, and adult duct cells. The following markers were used in determining the lineages, not both endocrine and exocrine cells derive from a common precursor. Neurog-3 - Neurogenin-3 (Ngn3) protein encoded in humans by the NEUROG3 gene. A basic helix-loop-helix (bHLH) transcription factor expressed in pancreas endocrine progenitor cells. This factor family involved in neural precursor cell determination in the neuroectoderm. CD133 - Prominin-1 a glycoprotein encoded in humans by the PROM1 gene. CD24 - Cluster of differentiation 24 or heat stable antigen CD24 (HSA) a protein encoded in humans by the CD24 gene. CD24 is a cell adhesion molecule. CD49f - Integrin alpha-6 (ITGA6) protein encoded in humans by the ITGA6 gene. Associates with a beta protein to form a laminin-binding heterodimers involved in adhesion.
, E15, E17, P1, P15, 8–12 week beta cells, P1 and 8–12 week alpha cells,")
139
Abnormalities Annular pancreas Annular pancreas - Annular pancreas is a distinct form of duodenal atresia in which the head of the pancreas forms a ring around the second portion of the duodenum. During the neonatal period, the clinical picture is dominated by epigastric distension with vomiting, which is nonbilious as the obstruction is usually supra-vaterian. Chromosomal abnormalities are present in one-third of cases of annular pancreas, with trisomy 21 (followed by trisomy 18 and 13) being the most frequently detected anomaly. Annular pancreas is an embryopathy resulting from an anomaly occurring early (towards the fourth week) in development. Occurs (1 in 7,000 people) where the pancreas forms as a "ring" of tissue surrounding the duodenum which is subsequently narrowed. Pancreas divisum Pancreas divisum - This is a congenital anomaly in the anatomy of the ducts of the pancreas in which a single pancreatic duct is not formed, but rather remains as two distinct dorsal and ventral ducts. Accessory pancreas Accessory pancreas - Accessory pancreas is an asymptomatic embryopathy characterized by the presence of pancreatic tissue in other sites of the body such as the splenic pedicle, gonadic pedicles, intestinal mesentery, duodenum wall, upper jejunum, or, more rarely, the gastric wall, ileum, gallbladder or spleen. Pancreatic tissue can also be located in Meckel's diverticulum. Agenesis-Aplasia of The Pancreas Agenesis-aplasia of pancreas - This refers to the failure of an organ to develop during embryonic growth and development due to the absence of primordial tissue of the pancreas.

140
Partial Agenesis Of Pancreas Partial agenesis of pancreas - Partial agenesis of the pancreas is characterized by the congenital absence of a critical mass of pancreatic tissue. The severity of the disease depends on the amount of functional pancreatic tissue present. Pancreatic agenesis is commonly associated with other malformations, in particular pancreaticobiliary duct anomalies, leading to acute or chronic pancreatitis, hyperglycemia (50% of cases), or, more rarely, polysplenia. Hypoplasia of Pancreas Diabetes Mellitus Maternal diabetes (and hyperglycaemia) have been shown to lead to increased fetal islet hyperplasia of the insulin producing beta cells and insulin secretion. Intrauterine Growth Restriction Intrauterine growth restriction can lead to a delayed development of the insulin producing beta cells and low insulin secretion. Tumours Serous Cystadenoma (endocrine tumour), Somatostatinoma (tumour of delta cell origin), intraductal papillary-mucinous neoplasm

148
The question of the origin and early development of the spleen has been the object of researches since the year A considerable amount of literature has accumulated since that time, in which the authors dealing with different classes of Vertebrata, and even those who have studied the question in the same animals, have arrived at widely differing conclusions. Some have believed the spleen to be derived from the mesenchyme of the dorsal mesentery, with or without the participation of the coelomic epithelium; some consider the coelomic epithelium to take the most important part in its development. Some, again, state that it arises together with the pancreas, while others believe it to have its origin in the endodermal epithelium of the alimentary canal. 1. The origin from the pancreas has been supported by Gotte in 1867 and Schenk in 1873, both of whom were dealing with birds. Kupffer in 1892 claimed to have found the spleen in ganoids arising as a diverticulum of the pancreas.

149
Woit in 1897 states that in birds and in Siredon it arises in genetic relation with the pancreas, but does not consider this to be the case with Rana. Glas (1900) describes a lieno-pancreas in Tropidonotus, with a duct leading into the intestine. Orru, Esio (1902), also gives the pancreas a share in the formation of the spleen. In this connection from Schenk His personal observations apply only to the chick and mammals. He considers that the spleen arises together with the pancreas, but is part of the mesoderm into which the pancreatic diverticulum grows. Kupffer, on the other hand (quoted by Laguesse), denitely gives the spleen an endodermal origin. In the Sturgeon he found four pancreatic buds, two dorsal and two ventral. The dorsal posterior diverticulum, according to him, divides into three branches: of these the right continues pancreatic in structure, the middle one forms the sub-chordal lymphoid tissue, while the left forms the spleen anlage. Toldt also states that in the human embryo the spleen begins by a thickening of the coelomic epithelium, which becomes cubical and splits into layers. Janosik points out the distinction between the mesenchyme and the mesothelium, and states that the proliferation of the latter forms the spleen anlage. This was particularly clear in Lacerta agilis. He nds no genetic relation between the spleen and the dorsal pancreas, though they are in close contact anatomically in Lacerta.
 describes a lieno-pancreas in Tropidonotus, with a duct leading into the intestine. Orr")
150
3. Various writers, from Peremeschko in 1867 (I) to Pinto in 1904, have affirmed that the spleen is mesodermal in origin. Laguesse deals very fully with the question of the formation and regeneration of blood corpuscles in the spleen in fishes, and considers its relation to the primitive sub-intestinal vein a very important one. Tonko, who deals in his paper entirely with the Amniota, concludes that the spleen in all these forms is entirely mesodermal in origin, but does not deny the possibility of the development being different in Fishes and Amphibia
 to Pinto in 1904, have affirmed that the spleen is mesodermal in origin. Laguesse deals very fully with the question of the formation and regeneration of blood corpuscles in the spleen in fishes, and c")
151
In contradistinction to J anbsik, Tonkoff does not make any essential distinction between the coelomic epithelium and the mesenchyme Pinto, to whose recent and detailed work is referred on several instances, believes the spleen to be of mesodermal origin, with the participation of the caalomic epithelium. 4. Maurer, whose conclusions differ widely from other authors, states that the spleen is of endodermal origin, and that it is formed by the mitosis of certain cells in the intestinal epithelium, which divide in a plane transverse to the surface of the intestine. The new cells are extruded into the surrounding mesenchyme, where they collect round the walls of small intestinal branches of the mesenteric artery. They then, according to Maurer, travel upwards in the walls of the arterial branches, until they arrive at the main trunk, and there collect to form the anlage of the spleen. He states that he found these cells, which he considers to be the first lymphoid elements, appearing around the small terminal branches of the arteries, before they could be traced at all on the main branches, or on the mesenteric trunk itself. He does not consider that any possible forerunners of lymphoid cells can be found in embryos of 46 mm. ano-buccal length (about 11 mm. total length), while in embryos of 6 mm. ano-buccal length he finds such forerunners only in the mesenchyme surrounding the intestine around the walls of the smallest terminal branches of the artery.

152
Choronschitzky, who studied the development of the spleen in Amphibia, reptiles and birds, agrees with Maurer that the intestinal epithelial cells migrate into the surrounding mesoderm, and give rise to mesenchyme tissues ; but he states that this occurs along the whole length of the intestinal tract, and not more in the region of the spleen than elsewhere. He draws attention to the part taken by the coelomic epithelium in the formation of the spleen Pinto does not deny Cl1nronschitzkys view, that the proliferation of the intestinal epithelium may cause cells to wander into the surrounding mesenchyme, though he regards it as being far from satisfactorily proved. If such migration does take place, it is as frequent in other parts of the alimentary canal as in the region of the spleen, and Pinto maintains that the extruded cells become mesenchymal elements. They cannot, therefore, be regarded as forming an endodermal origin for the spleen. He is unable to conrm those subtle distinctions which Maurer makes as to the varying directions of the equatorial planes of mitoses which are to be seen in the intestinal epithelium. Here a summary is given in tabular form of the views taken by the different investigators as to the origin of the spleen.

154
In The Stage I Embryo 10 mm. total length, 4 mm. ano-buccal. Stained with Methyl Blue A Transverse sections of 10,u. Liver and pancreas well developed, Walls of intestine thick and convoluted. Many granulated cells containing yolk still to be found in them.

155
It will be seen from the above table that the majority of writers have taken the view that the spleen is developed from the mesenchyme, and it might seem unnecessary to go once more into the question, but it must be remembered that many of the authors quoted have only touched supercially on the question, and have dealt with the development of the spleen in different classes of vertebrates. Tonkoffs important work deals only with the Ainniota, and, in spite of Pintos comprehensive and careful study of the question in Amphibia, Reptiles, Birds and Mammals, there still remains the fact that Maurer, and to some extent Choronschitzky, hold views which are widely different from those of other authors. It seemed worth while then, as the question, for Amphihia at any rate, is still open, to try and arrive at denite conclusions, and therefore a series of embryos of Rana temporaria was obtained. In the following account the most important details of the various stages are described.

156
Descriptive Embryology These series of specimens include embryos of all stages from 10 mm. total length (4 mm. anobuccal) to 27 mm. total length (11 mm. ano-buccal). (In older stages, the total length is no guide to the age of the embryo, and the ano-buccal length is not always dependable when the development of the limbs has begun.) Both longitudinal and transverse sections were made, but the latter was found to be more useful. Most of the sections were stained in bulk with Haematoxylin and Eosin. In an embryo of 27 mm. total length, in which the limbs have not yet appeared externally, the spleen appears as an oval body of smooth outline and of considerable size. It lies partly anterior to and partly on the left side of the mesenteric artery, in close contact with its wall, and appears in transverse sections as if wedged between the artery and the intestine which lies on its left, and to which it is attached by mesentery. It is composed of denite lymphoid tissue and contains blood-spaces, but is very uniform in its appearance. It is quite clearly separated from the artery and the intestine, and lies in the mesentery surrounded by a distinct limiting membrane. The spleen is comparatively late in developing and its growth is slow. Even when the limbs are well developed, the spleen shows very little advance on the condition to be seen in an embryo of 18 mm. total length.
 to 27 mm. total length (11 mm. ano-buccal). (In older stages, the total length is no guide to the age of the embryo, and the ano")
158
Mesenteric artery well developed, sending branches to pancreas and alimentary canal. The walls of the mesenteric artery are surrounded throughout by a thick coat of round cells with large nuclei (PLY). These are to be seen both on the branches and on the main trunk, and already a small heaping up of such cells may be seen on the left of the mesenteric artery, exactly in the position which the spleen will later on occupy (SPA). The cells, although not unlike the cells of the intestinal epithelium, are not identical with them. Mitosis is certainly going on in the intestinal epithelium, but only a few of the epithelial cells show any indication of being squeezed out into the surrounding mesoderm (See Legend Figure Below).
. These are to be seen both on the branches and on")
160
That this epithelial extrusion is not limited to early stages, nor to that part of the alimentary canal in the neighbourhood of the spleen, is shown in the Legend figure below, which shows such extrusion taking place. The section was taken through a part of the canal remote from the spleen, and at a stage when the spleen was already a denite" structure, covered by a limiting membrane. In embryos of 10 mm total length, Where the spleen is in process of formation, the intestinal Walls are not unstratied as in Maurers figure, but are thick and convoluted, which makes it extremely difcult to determine in what plane mitosis takes place (fig. 2, I). The epithelium is clearly divided in all situations from the mesenchyme lying outside it, and the round nucleated cells which are to be seen in the mesoderm among the more spindle-shaped cells are smaller and more granulated than the epithelial cells of the intestine, and. resemble more closely the cells surrounding the Walls of the arteries (fig. 2, PLY). Again, many'of these last-mentioned cells are undergoing mitotic division, although Maurer states that no such division could be seen in the connectivetissue cells at a similar stage. They are certainly forerunners of lymphoid cells, and are, even at this stage, grouped as thickly around the mesenteric artery itself as round its terminal branches, even although this is an earlier stage than any described by Maurer, and there is no indication that these cells are descendants of migrants from the intestinal epithelium.

161
It may be objected that the cells above mentioned are not denitely lymphoid in character; but the condition I have described, and which is shown in fig. 2, persists, and the spleen is developed by proliferation of the cells, which gradually become more lymphoid in character, and surround the dorsal part of the artery at the level at which the organ is situated when it is completely established. Exactly the same kind of cells are found, as Maurer himself says, in the connective tissue of the kidney, the genital gland, etc., which could not have travelled by the same path as those followed, according to him, by the spleen cells. In other embryos of 10 mm. total length the thickening of the mesenchyme cells is more general, and no denite spleen anlage can be distinguished.

162
Stage II Embryo 15 mm, stained methyl blue. Walls of intestine thinner and more simple. Single-layered epithelium shows distinct rounded body on the left side of the mesenteric artery. It is attached by mesentery to the part of the intestine lying to the left of it, and lies close against the wall of the artery. It extends through seven sections of 5u, and is then lost as a lymphoid thickening on the wall of the mesenteric artery. It is covered by a single layer of coelomic epithelium, which is continuous With that reflected over the intestine and the body Wall. It is in very close contact with the arterial Wall, but is distinct from it. The cells are for the most part rounded, granular, with darkly stained round nuclei. They are in process of cell division, which can be seen in various stages. The tissue also contains blood corpuscles. There are no blood-spaces. In some embryos even of slightly greater length the spleen is less denite. The lymphoid tissue is present in large quantity, and is heaped up around the artery at the level Where the spleen eventually comes to lie. In one series of sagittal sections the splenic thickness is very. Well marked, and the mesenteric artery passes through its posterior part. This is probably due to the fact that the spleen is closely associated with the lymphoid tissue surrounding the wall of the artery, and occasionally extends backwards into closer connection With it. The same remarks concerning the character of the cells apply as in the last stage described. Scattered cells are to be seen which appear to be identical with the endodermal epithelial cells, and may have been extruded in the process of cell division, but they are quite casually distributed, and the difference between them and the lymphoid cells is clearly marked (See Legend figure Below).

163
Embryo 18 mm total length. Stained haematoxylin and eosin. The spleen runs through nine sections of.10. It is considerably more denite than in the last stage, and lies freely in the mesentery, though in very close contact with the wall of the artery, which is still surrounded by scattered lymphoid cells, and with the part of the intestine which lies to its right. The coelomic epithelium covers the free surface, but there is not a denite capsule (fig. 1, SP). The cells composing it are already denitely lymphoid in character (fig. 3, LY). As compared with those of the intestinal epithelium, they differ markedly from the latter, being smaller, more irregular, and inclined to be elongated in shape and more granular, the. protoplasm taking a deeper stain, While the nucleus is more diffuse. At this stage mitosis is still proceeding in the cells of the intestinal epithelium, but it is diicult even yet to determine the plane of division on which Maurer lays so much stress (fig. 3, IE). Here and there, cells derived from these mitotic divisions lie outside the basement membrane in the mesenchyme surrounding the intestinal wall. They are also to be seen here and there along the walls of branches of the mesenteric artery, but they do not collect specially round its terminal branches, as Maurer asserts, and they appear to be quite distinct from the lymphoid cells on the arterial Wall, which are to be seen in the same sections. A certain amount of emigration from the intestinal epithelium does take place, but there seems no reason for believing that the cells which have migrated take any direct part in the formation of the spleen.

164
Stage III Embryo 24 mm. Sagittal section. Stained Haematoxylin and Eosin. The spleen runs through eighteen sections of 10,u.; it lies against the wall of the mesenteric artery, to the left side of and anterior to it. It is surrounded by a capsule of attened epithelium, and is separated from the artery wall. It is dorsal and posterior to the stomach. The cells are granulated, the majority are rather large and rounded, and the epithelial capsule is incomplete. Various stages of cell division can be seen in the splenic cells, many blood corpuscles are scattered through the tissue, and some dark elongated cells are present, which often divide transversely.

166
Embryo 27 mm. total length, 11 mm. ano-buccal. Stained haematoxylin and eosin (g. 4). Shows the spleen as an oval, smooth body, running through thirtyfive sections of 10. It lies freely in the dorsal mesentery, by the side of the mesenteric artery. At its greatest diameter it is in very close contact with the artery, and with the intestine to the left of it, but it is separated from it, both by the coelomic epithelium and by its own distinct limiting membrane. In some sections a small blood-vessel is seen running between the intestine and the spleen, and sending branches into it. The dorsal mesentery is very diffuse and thin, and shows many sections through blood-vessels. Cells consist of (1) large cells with granular nuclei in various stages of cell division ; some are rounded, others spindle shaped or oval. (2) Smaller elongated cells, very deeply stained; these are possibly blood corpuscles in an imperfect state. (3) Red blood corpuscles (see Legend Figure Below). In some sections small vessels can actually be seen entering the spleen tissue. There are no denite Malpighian corpuscles, but the Whole tissue is permeated with blood corpuscles.
. Shows the spleen as an oval, smooth body, running through thirtyfive sections of 10. It lies freely in the dorsal mesentery, by the side of the mesenteric artery. A")
168
In an embryo in which the limbs were Well developed and the tail partly absorbed, the spleen shows Very little further advance in structure, but the capsule -is rather more developed. It Will be seen from the Legend figure immediately above that the lymphoid tissue which surrounds the artery is less dense than in earlier stages of development, and is more conspicuous around the main trunk of the artery than at its terminal branches. The lymphoid mesenchyme cells (LY) in the Legend figure immediately above, ) around the artery are exactly similar to those in the spleen itself, and differ markedly from the cells which appear to be. strayed endodermal cells from the epithelium of the alimentary canal. In the Legend Figure immediately above, there are a few of these cells (IE) to be seen lying in the mesentery among the mesenchyme cells. It will be seen that they differ in appearance from the latter, While they bear a strong resemblance to the cells of the intestinal epithelium, being large, clear, and spherical in shape, with very denite nuclei.

170
In the same series of sections, similar cells are to be seen lying in the mesentery in parts of the alimentary canal remote from the spleen, and they appear to be quite impartially, though sparsely, distributed. The first possible forerunners of the lymphoid cells of the spleen are then to be found in embryos of 10 mm. total Anobuccal),length of (about 4 mm). The mesenteric artery at this stage has a thick coat of mesenchyme cells, which have begun to be heaped up in the position to be occupied by the spleen. These cells are grouped thickly round the main trunk and branches of the mesenteric artery, and in no instance was it found that such cells surround the terminal branches, and not the artery itself. These cells differed in appearance from those of the intestinal epithelium in size, shape, and appearance of nucleus. Cells identical With the intestinal epithelium were to be seen here and there in the mesentery, but were few in number, and easily to be distinguished from the mesenchyme cells among which they lay (see Legend figure immediately above). In no case were there such cells in the spleen anlage itself, nor was there any transition stage which could lead me to suppose that they were altering their character. The appearance of the above mentioned cells, both in the mesentery and in the mesenchyme surrounding the intestine, conrm

171
Maurers View that there is a migration of intestinal epithelium, but the migration takes place along the whole length of the intestinal canal, and continues long after the spleen is a distinct structure and the cells take no part in spleen formation. The cells were more frequent in the later stages than in the less developed embryos, and even at the stage at which Maurer first notes their appearance the spleen anlage has begun to appear. Even before there is any trace of the actual spleen, the trunk of the mesenteric artery shows the thick coat of mesenchyme cells, which are undoubtedly primitive lymphoid tissue. The coelomic epithelium certainly forms the capsule, but it does not appear to take a larger share than this in the development of the spleen. The limiting membrane is very distinct from the rest of the tissue throughout its development.

172
Conclusions In the first place, it may be stated that the pancreas, in The Rodent at least, can have no part in the formation of the spleen. The two organs are not in contact at any stage in the development of either, and the pancreas is already well developed when the splenic Anlage first appears. Secondly, for reasons which it is hoped have been made clear in the last section, I do not believe that the endodermal epithelium of the intestine is responsible for the formation of the spleen, even indirectly. The migration of cells from the intestinal epithelium, where it does occur, takes place at a stage when the spleen Anlage is already formed, and when the cells of which it is composed are themselves proliferating rapidly. However, that these epithelial cells do go to increase the mesodermal tissue of the mesentery seems clear, though it is speculative that the part they play is a very considerable one. The conclusion then is, that the spleen in Mammalian species arises from the mesenchyme tissue of the dorsal mesentery, in close connection with the mesenteric artery, as a thickening of the lymphoid tissue which surrounds the wall of the artery at an early stage. It develops by proliferation and differentiation of these primitive lymphoid cells, and becomes highly vascular. The coelomic epithelium appears to form the capsule, and perhaps enters into the formation of the reticular network.

Еще похожие презентации в нашем архиве:







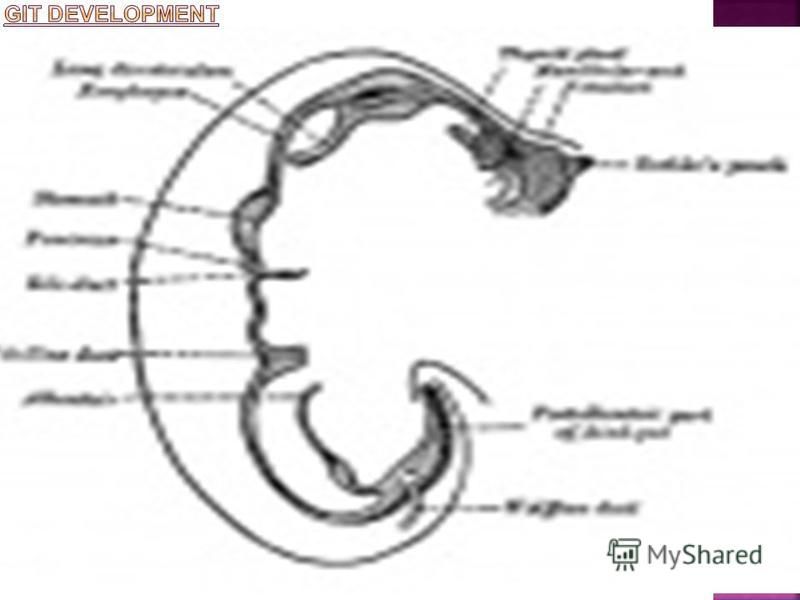











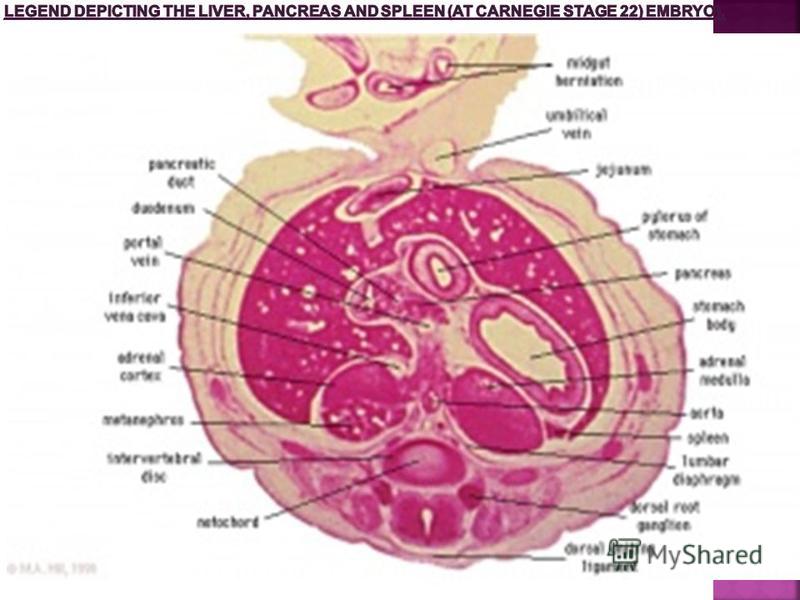






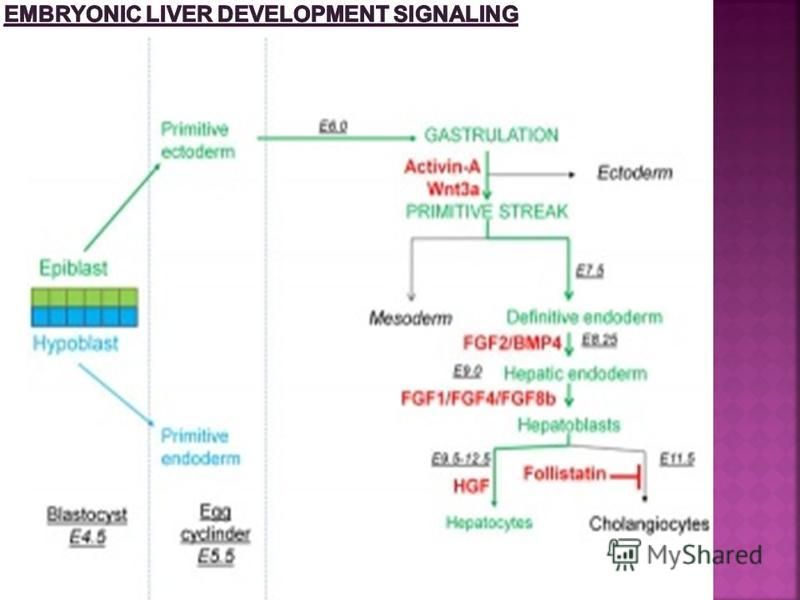



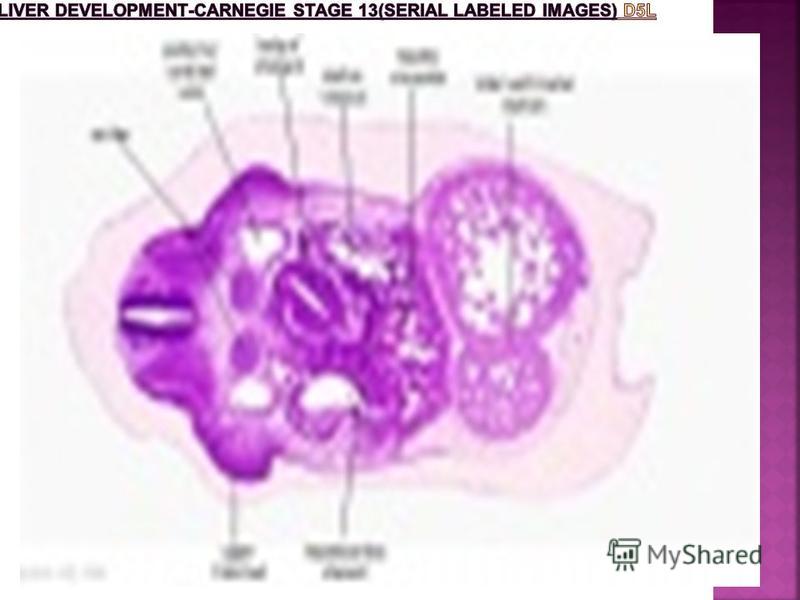



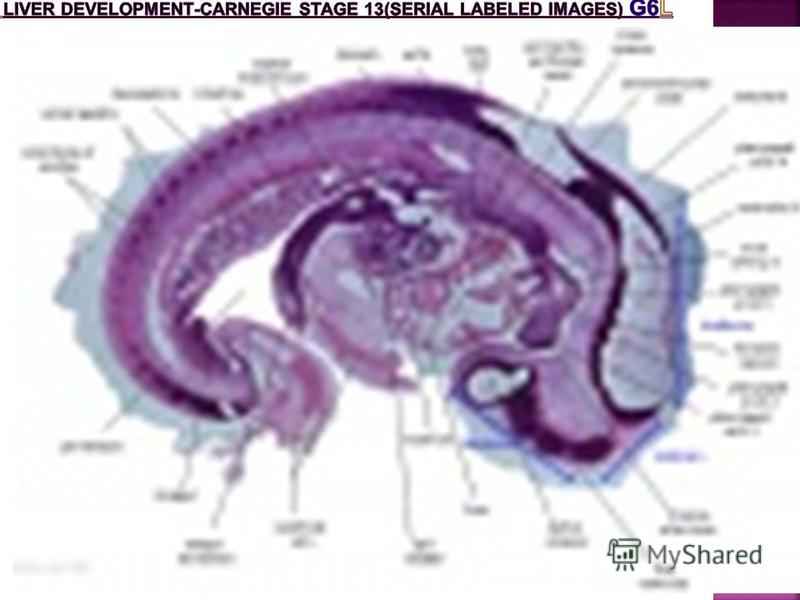



























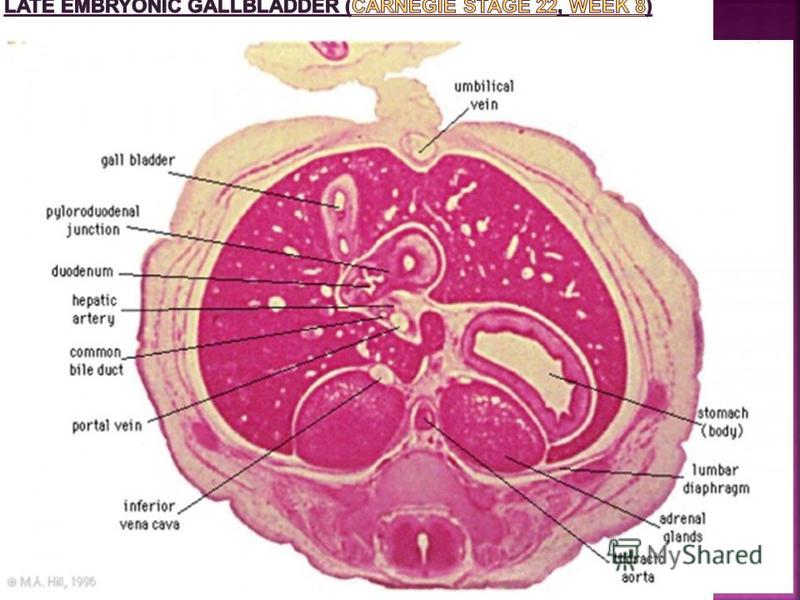


























. Paraganglia.")
















