Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Мавзу: Оқсиллар биосинтези Мавзу: Оқсиллар биосинтези

2
Кўриб чиқиладиган саволлар Генетик код ва унинг таркиби. Ниренберг ва Маттей тажрибаларининг генетик кодни тасдиқлашдаги аҳамияти. Оқсил биосинтези босқичлари. Посттрансляцион ўзгаришлар. Оқсил биосинтезининг бошқарилиши. Оқсил биосинтези ингибиторлари. Генлар таъсирининг бошқарилиши ва хужайра дифференцировкаси. Молекуляр генетика асослари. Наслий касалликларнинг молекуляр механизми. ДНК шикастланиши ва репарация. Оқсиллар полиморфизми. Тўқима номутаносиблиги

3
Генетик код

4
Генетик коднинг хусусиятлари: Биологик код триплет ҳисобланади. Бир аминокислота учун бир неча код бўлади, фақат метионин ва триптофанни коди битта. Код узлуксиздир Код универсал характерга эга. Ҳаммаси бўлиб 64 та триплет код бўлиб, 61таси 20 хил аминокислотани кодлайди, қолган 3таси- УГА, УАА, УАГ –маъносиз (нонсенс) триплет бўлиб, бирорта аминокислотани кодлаштира олмайди. Улар трансляцияни чегаралаш функциясини бажаради, шу сабабли стоп-кодонлар деб аталади.

5
Комплементарлик принципи нуклеотидлар учун ҳарактерли ҳисобланади, лекин нуклеотидлар ва аминокислоталар ўзаро комплементар бўла олмайди. Шунинг учун аминокислоталар кодонлар ёрдамида полипептид занжирга тўғридан-тўғри бирика олмайди. Аминокислоталарни мРНКнинг маълум участкасига бириктириш адапторлар ёрдамида юзага келади ва бу вазифани тРНК бажаради. Аминокислота тРНКга бирикиб, ўзининг триплет коди билан бирикиш хоссасига эга бўлади.

6
Оксил синтезининг асосий босқичлари: Аминокислоталарнинг фаолланиши Полипептид занжир инициацияси ЭлонгацияТерминация Ўз-ўзидан ўралиш ва процессинг

10
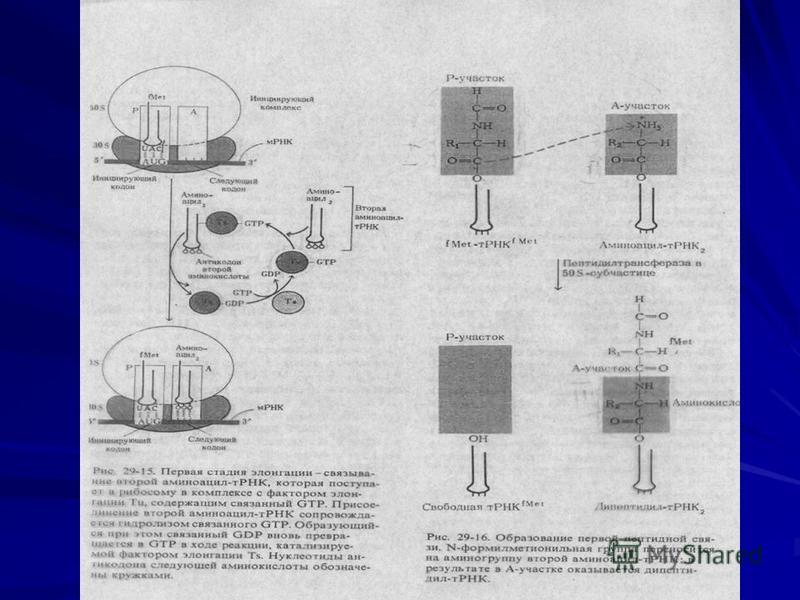
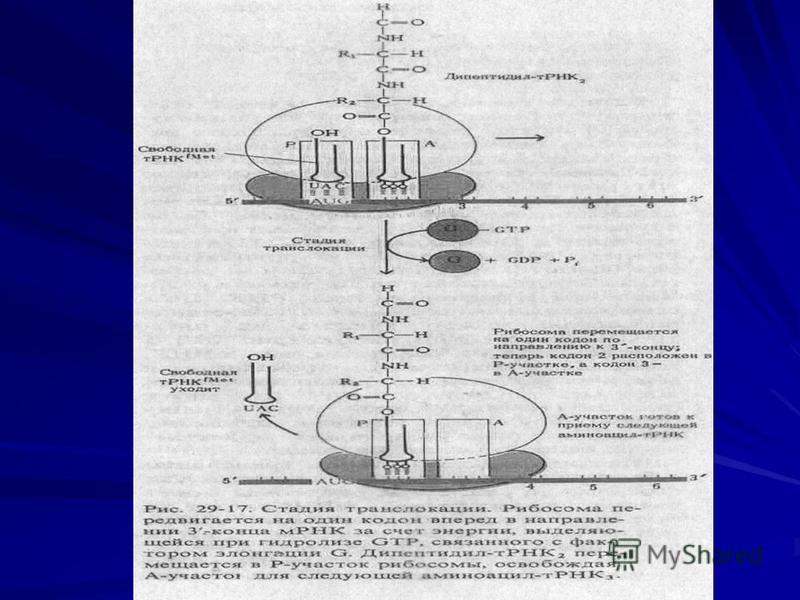
Элонгация

11
Терминация кодонлари УАА, УАГ, УГА УАА, УАГ, УГА Терминация босқичида қатнашувчи омиллар: R1, R2, R3

12
Пептидил-тРНКПептидил-тРНК 3' Пептидил-тРНК Свободный полипептид 3 3

13
Полипептид занжирнинг посттрансляцион модификацияси Кўпчилик модификациялар ЭР фолдинг асосида полипептид занжирини шаклланишида кечади. У қуйидагиларни ўз ичига олади: Қисман протеолиз Ковалент модификация (фосфорилланиш, гликирланиш, ацетилланиш) Қўшимча моддаларни киритилиши (простетик гуруҳлар, коферментлар, металлар, гидроксил гуруҳлар)
 Қ")
14
E.Coli Lac-оперонининг экспрессиясининг бошқарилиши

15
Эукариотларда генлар экспрессияси бошқарилиш механизми Структур генлар миқдорини ўзгариши Хромосомаларда генларнинг алмашинуви Геномнинг турли хил қисмлардаги транскрипциянинг самарадорлиги билан Бирламчи транскиптонни модификацияси Трансляция даражасида (энхансерлар ва сайленсерлар таъсирида) Синтезланган полипептид занжирининг посттрансляцион ўзгариши билан

17
Нуклеин кислоталар ва оксил синтезининг антибиотиклар билан ингибирланиши: Пурамицин – элонгацияни тўхтатади Актиномицин Д – РНК полимеразани ингибирлайди Рифамицин – РНК полимеразани ингибирлайди Хлорамфеникол – пептидилтрансферазани ингибирлайди Циклогексамид – транслоказани ингибирлайди Стрептомицин, неомицин – транслоказа хатоликларини вужудга келтиради Эритромицин, олеандомицин – транслоказа фаоллигини тормозлайди.

18
Мутациялар Генда пурин ва пиримидин асослари кетма- кетлигини репарация ферментлари таъсирида тикланмаслиги мутация дейилади. Тахминан 20% ҳомиладорликда эмбрион хромосомасида структур ўзгаришлар кузатилади. Уларнинг 90% ҳомила ривожланишини бузилишига ва спонтан абортга олиб келади, айниқса биринчи ҳафталарида.

19
Мутациялар таснифи Мутация тури Мутацион ўзгаришлар мисоллар Геном Хромосомалар сонини ўзгариши Даун касаллиги (қўшимча хромосома) Хромосом Хромосомаларнинг сони ўзгармаган ҳолда уларнинг тузилишини ўзгариши Дюшеннинг мушак дистрофияси (Х хромосоманинг делецияси) Ген Кодон ёки геннинг маълум бир қисмини ўзгариши Глобиннинг β-занжири генида 1 нуклеотидни ўзгариши натижасида келиб чиқадиган ўроқсимон анемия
 Хромосом Хромосомаларнинг сони ўзгармаган ҳолда уларнинг тузилишини ўзгариши Дюшеннинг мушак дистрофияси (Х хромосомани")
20
Ген мутациялар Кодонда нуклеотидни алмашинуви (миссенс – қўшимча, нонсенс – аминокислотани бўлмаслиги) мутация. Кодон мазмуни кўпам ўзгармайди, чунки генетик код айнаган. Делеция – полинуклеотид занжирида мономерни еки триплетни тушиб қолиши. Кодон ўқилиши силжийди ва бошқа оқсил синтезланади. Қўшимча мономерни киритилиши. Бунда битта еки бир-неча аминокислотага узайган оқсил синтезланади ва функцияси йўқолади. Мутациялар нейтрал, ижобий ва салбий бўлади. Жинсий хужайралардаги мутациялар наслий касалликларга олиб келади. Соматик хужайралардаги мутациялар функционал ўзгаришлар ва баъзи ҳолларда ўсмалар келиб чиқади. Агар мутация натижасида оқсил ижобий хусусиятга эга бўлса, организмнинг мослашуви кучаяди. Улар табиий танлашни таъминлайди.
 мутация. Кодон мазмуни кўпам ўзгармайди, чунки генетик код айнаган. Делеция – полинуклеотид занжирида мономерни еки триплетни тушиб қолиши. Кодон ў")
21
Наслий касалликлар Бирламчи мутацияларда улар гетерозигота ҳолатда бўлиб, касалликни намоён қилмайди, аммо популяцияда тарқалишига сабаб бўлади. Мутациялар ота ва онада бўлса улардан ҳосил бўладиган авлодда гомозигота ҳолат кузатилиб касаллик ривожланади. 2-4% туғилган чақалоқларда наслий касалликлар кузатилади, ўлим ва ёшликдан ногиронликнинг 40% сабаби бўлади. Ген мутацияларининг 31% ферментларга тўғри келади. Шу билан бирга баъзи калассикларга оилавий наслий моиллик бор. Масалан, атеросклероз, қандли диабет, оилавий полипоз, подагра, шизофрения ва бошқалар.

22
Оқсиллар полиморфизми Ҳар қандай ген маълум даражада ўзгарувчанлик билан тавсифланади. Филогенезда геномнинг мураккабланиши 2 йўналишда кечади: 2 маротаба ортиши ва улардаги мутациялар. уларнинг популяцияда тарқалиши фенотипик гетерогенликга ва оқсиллар полиморфизмига, яъни бир хил ёки ўхшаш функцияларни бажарувчи оқсилларни ҳосил бўлишига. 2 турдаги оқсиллар полиморфизми мавжуд: Геномда локусларнинг ўхшашлиги натижасида ҳосил бўлган ноаллел генлар (HbA, HbA2, HbF). Кўп аллел генлар (HbA, HbS).

23
Иммуноглобулинлар 5 синф иммуноглобулинлар мавжуд:A, B, D, E, M

24
Қон гуруҳлари Эритроцитлар антигени Йўқ АВАВ Генотип00 АА ёки А0 ВВ ёки В0 АВ Қон зардобидаги антителолар Анти-А ва анти-В Анти-ВАнти-Айук Қон гуруҳи 0 (I) А (II) В (III) АВ (IV) Учраши (%)
 А (II) В (III) АВ (IV) Учраши (%) 4540105")
25
Тўқима номутаносиблиги Тур ичи спецификлигини таъминловчи ва тўқима номутаносиблигига олиб келувчи оқсиллардан бири HLA (одам лимфоцити антигени). Улар 2 синфга бўлинади: I синф – гетеродимер оқсиллар, барча ядро тутувчи хужайраларда мавжуд. Унинг α 1 ва α 2 доменлари хужайра юзасида жойлашган антигенларни танийди. II синф – гетеродимер оқсиллар, В-хужайралар, Т- хелперларда мавжуд, Т-киллер ва макрофагларда бўлмайди. Уларнинг α- ва β-занжирлардаги доменлари ангиген билан боғланади. Улар жуда юқори полиморфизмга эга ва хужайра номутаносиблигини белгилайди. Оқсиллар полиморфизми шунчалар юқорики, ҳар бир инсонни биокимёвий индивидуаллигини белгилайди.
. Улар 2 синфга бўлинади: I синф – гетеродимер оқсиллар, барча ядро тутувчи хужайраларда мавжуд. Унинг α")
26
Ген инженерияси ДНК-полимеризация реакцияларини тиббиётда қўлланилиши (ПЦР-диагностика). Турли хил организмлар ДНК сининг бирламчи қурилиши турлича. Касаллик чақирувчи омиллар ДНК сига хос праймер танлаб реакция ўтказилганда праймер қўзғатувчининг ДНК си билан гибридизацияланади. Масалан, ПЦР-диагностика гепатитнинг В ва С лари борлигини ва бошқа турли хил инфекцияларни аниқлаш имконини яратади. Инсон геномини ўрганиш. Бунда ҳам махсус праймерлардан фойдаланилади. Бу усулда турли наслий касалликларни аниқлаш мумкин. Масалан, филадельфия хромосомаси сурункали миелолейкозда мавжудлигини, оилавий полипозни келтириб чиқарувчи оқсилни ва бошқалар.
. Турли хил организмлар ДНК сининг бирламчи қурилиши турлича. Касаллик чақирувчи омиллар ДНК сига хос праймер танлаб реакция ўтказилганда праймер қўзғатувчининг ДН")
27
Ген инженерия усулида E.Coli трансформацияланган хужайраларидан инсон препроинсулинини синтезлаш

Еще похожие презентации в нашем архиве:



 проекциялар Мақсад: Геометрик проекцияларни, хусусан марказий проекцияларни ўрганиш. Калит сўзлари: Геометрик проекциялар,")
")


 (www.gov.uz)")




лар қўйиш орқали баҳолашингиз, ўз видеоингизни кўчириб, дўстларингиз.")









