Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
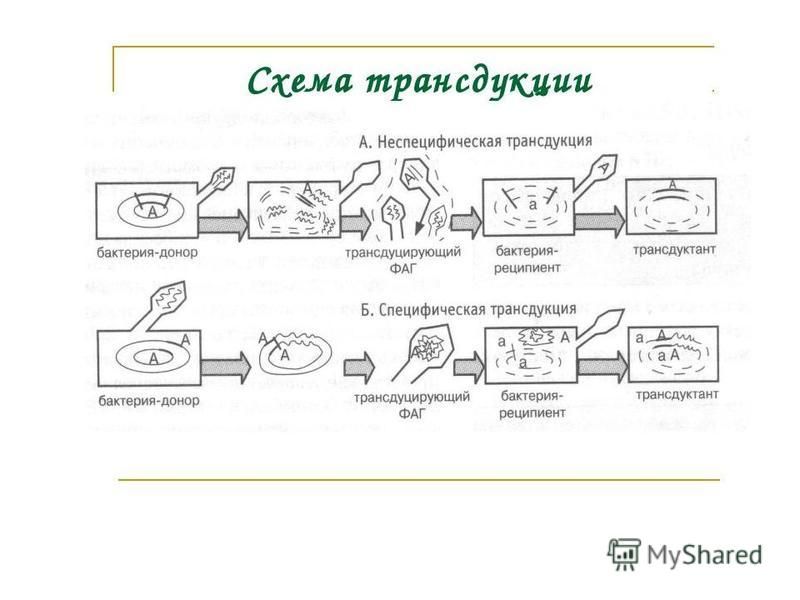
6. Трансдукция – перенос генетического материала из клетки донора в клетку реципиента через трансдуцирующий бактериофаг. Последний представляет собой умеренный фаг, который в состоянии профага получил участок ДНК от донорской клетки в результате неточного вырезания своей последовательности из генома клетки-донора. При этом бактериофаг становится дефектным, т.к. теряет часть собственной нуклеиновой кислоты. Такой фаг упаковывается в свою оболочку, выделяется из клетки и может проникать в клетку-реципиент. Этот вид рекомбинаций открыт Н. Циндером и Дж. Ледербергом в 1951 г. Различают 3 вида трансдукции: 1. Неспецифическая; 2. Специфическая; 3. Абортивная.

2
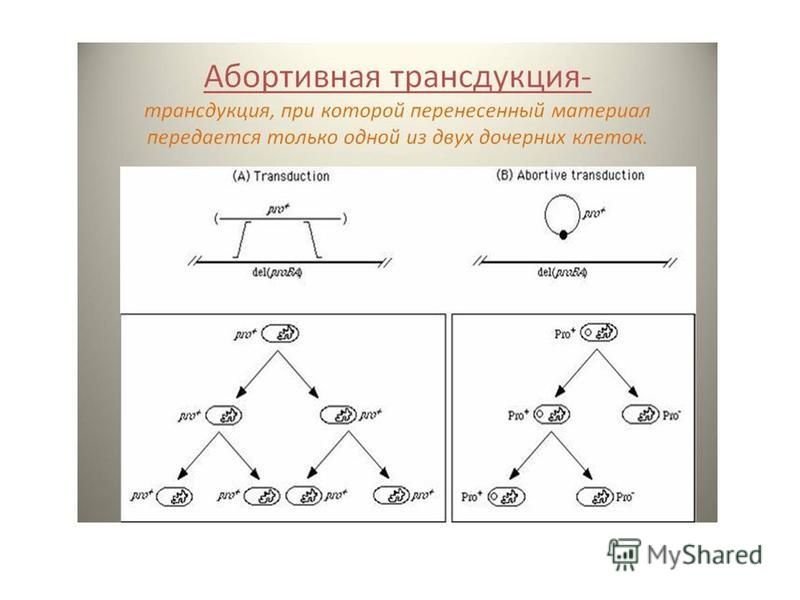
Неспецифическая трансдукция. При этом трансдуцирующий бактериофаг передает в реципиентную клетку любой ген донорской клетки и включает его в гомологичную область ДНК реципиента путем рекомбинации этого гена с нуклеоидом. Трансдуцирующий бактериофаг выступает лишь в роли переносчика, в нуклеоид не встраивается, и лизогенизации реципиентной культуры не происходит. Специфическая трансдукция. Здесь бактериофаг переносит строго определенный ген (или гены) от клетки донора к реципиенту и встраивает его в определенном участке ДНК реципиента путем сайт-специфической рекомбинации. В этом случае бактериофаг может встраиваться в нуклеоид клетки-реципиента, т.е. происходит лизогенизация бактерии. При этом такие клетки становятся невосприимчивыми, как и все лизогенные клетки, к последующему заражению гомологичным вирулентныйййм фагом. Обычно при специфической трансдукции переносятся бактериальные гены, сцепленные с геномом встроенного бактериофага. Чаще всего они окаймляют (фланкируют) профаг. Для E.coli и фага лямбда это гены gal и bio, контролирующие, соответственно, метаболизм галактозы и синтез витамина биотина. Абортивная трансдукция. В этом случае фрагмент ДНК донора, доставленный при трансдукции, не включается в ДНК реципиента и остается в цитоплазме. Клетка не лизогенизируется, а новый признак по мере деления клетки исчезает.

5
7.

6
8. Плазмиды внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой ДНК. По размерам составляют 0,15 % ДНК хромосомы. Плазмиды несут гены, не обязательные для клетки-хозяина, придают бактериам дополнительные свойства, которые в определенных условиях окружающей среды обеспечивают их временные преимущества по сравнению с бесплазмидными бактериами.

7
Плазмиды образованы молекулами ДНК. Регуляторные плазмиды участвуют в компенсировании тех или иных дефектов метаболизма бактериальной клетки. Кодирующие плазмиды привносят в бактериальную клетку новую генетическую информацию, кодирующую новые, необычные свойства (например, устойчивость к антибиотикам) ГРУППЫ ПЛАЗМИД F-плазмиды контролируют синтез F-пилей, способствующих передачи генетического материала от бактерий-доноров (F+) к бактериам- реципиентам (F–) в процессе конъюгации R-плазмиды (от англ. resistance, устойчивость) кодируют устойчивость к лекарственным препаратам. Плазмиды патогенности контролируют вирулентныйййе свойства бактерий и токсинообразование (плазмиды включают tox+-гены). Плазмиды бактериоциногении кодируют синтез бактериоцинов - белковых продуктов, вызывающих гибель бактерий того же или близких видов

8
Плазмиды могут определять вирулентность бактерий, например возбудителей чумы, столбняка, способность почвенных бактерий использовать необычные источники углерода, контролировать синтез белковых антибиотикоподобных веществ бактериоцинов, детерминируемых плазмидами бактериоциногении, и т. д. Существование множества других плазмид у микроорганизмов позволяет полагать, что аналогичные структуры широко распространены у самых разнообразных микроорганизмов.

10
10. Генетическая инженер́риа (генная инженерриа) совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами, введения их в другие организмы и выращивания искусственных организмов после удаления выбранных генов из ДНК[1]. Генетическая инженерриа не является наукой в широком смысле, но является инструментом биотехнологии, используя методы таких биологических наук, как молекулярная и клеточная биология, генетика, микробиология, вирусология.
 совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами, введения их в другие организмы и выращивания ис")
11
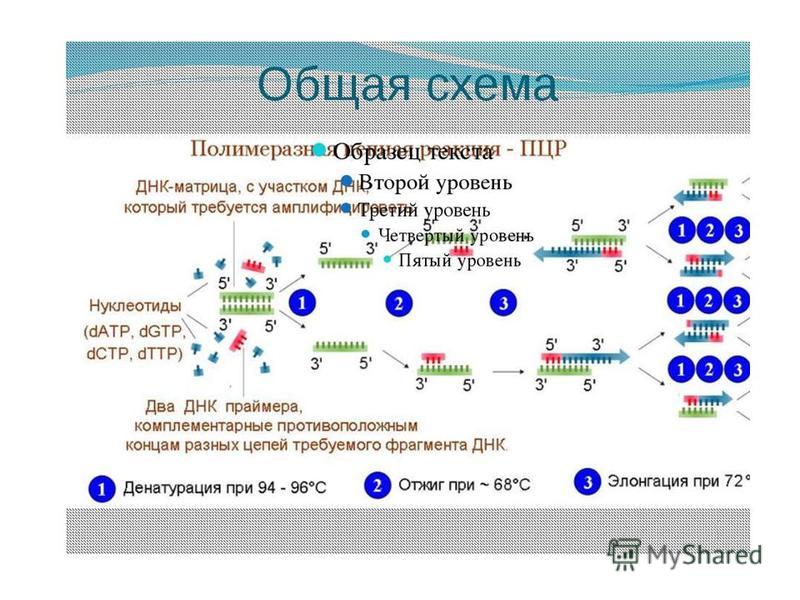
11.Полимера́зная цепная́я реа́кция (ПЦР) экспериментальный метод молекулярной биологии, позволяющий добиться значительного увеличения малых концентраций определенных фрагментов нуклеиновой кислоты (ДНК) в биологическом материале (пробе) Метод основан на многократном избирательном копировании определенного участка ДНК при помощи фермента Taq- ДНК-полимеразы. Полимеразная цепнаяя реакция позволяет получить амплификаты длиной до нескольких тысяч пар нуклеотидов. Для увеличения длины ПЦР-продукта до тыс. пар нуклеотидов применяют смесь различных полимераз, но все равно это значительно меньше длины хромосомной ДНК эукариотической клетки. Реакция проводится в программируемом термостате (амплификаторе) - приборе, который может проводить достаточно быстро охлаждение и нагревание пробирок (обычно с точностью не менее 0,1 °С). Амплификаторы позволяют задавать сложные программы, в том числе с возможностью «горячего старта» и последующего хранения. Для ПЦР в режиме реального времени выпускают приборы, оборудованные флуоресцентным детектором. Существуют также приборы с автоматической крышкой и отделением для микропланшет, что позволяет встраивать их в автоматизированные системы. Обычно при проведении ПЦР выполняется циклов, каждый из которых состоит из трех стадий: денатурации, отжига праймеров, элонгации (рис. 6.1 и 6.2). На рис. 6.1 представлена динамика изменения температуры в пробирке при проведении цикла ПЦР.
 экспериментальный метод молекулярной биологии, позволяющий добиться значительного увеличения малых концентраций определенных фрагментов нуклеиновой кислоты (ДНК) в биологическом материале (пробе) Метод основан")
13
12. Диссоциация – это особый, присущий только бактериам вид изменчивости, при котором происходит расщепление в пределах одного вида на S- и R-формы микроорганизмов. Это явление впервые исследовали Э.Вейль и А.Феликс (1917 г.). В основу этого подразделения положены генетические перестройки, приводящие к изменению ряда свойств (культуральных, антигенных, биохимических). Так, S-формы (англ. smooth – гладкий) чаще вирулентныййй, обладают хорошо выраженными антигенными свойствами, имеют капсулу, на средах дают рост мелких блестящих колоний. R-формы (англ. rough – грубый, неровный) реже вирулентныййй, не имеют капсулы, колонии крупные, шероховатые. Однако не у всех микробов S-форма свидетельствует о вирулентности. Так сибиреязвенные культуры, возбудители туберкулеза и чумы вирулентныййй в R-форме. Диссоциация обычно протекает в одном направлении: от S- к R-форме, иногда через промежуточные стадии образования слизистых колоний. Причиной диссоциации могут быть мутации, возникающие после встраивания внехромосомных факторов наследственности (эписом и умеренных фагов) в нуклеоид. Мутации сопровождают и процессы встраивания в нуклеоид транспозонов и инсерционных последовательностей. Если эти мутации нарушают функцию оперонов, которые отвечают за образование липополисахаридов клеточной стенки микроба, то образуются R-формы. Они дают шероховатые колонии, меняют антигенные свойства и ослабляют патогенность. Тем не менее, у дифтерийных бактерий S-R- диссоциация связана с их лизогенизацией бактериофагом; при этом R-формы образуют токсин, и их вирулентность резко увеличивается. Значение диссоциации заключается в получении бактериами селективных преимуществ, обеспечивающих их существование в организме человека или во внешней среде. Например, S-формы более устойчивы к фагоцитозу. R-формы, в свою очередь, более устойчивы к факторам окружающей среды.
. В основу этого подразде")
Еще похожие презентации в нашем архиве:







 совокупность приёмов, методов и технологий получения рекомбинантных.")

 совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК,")



 совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма.")








