Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
ЭРИТРОПОЭТИН Астана, 2016 г.

2
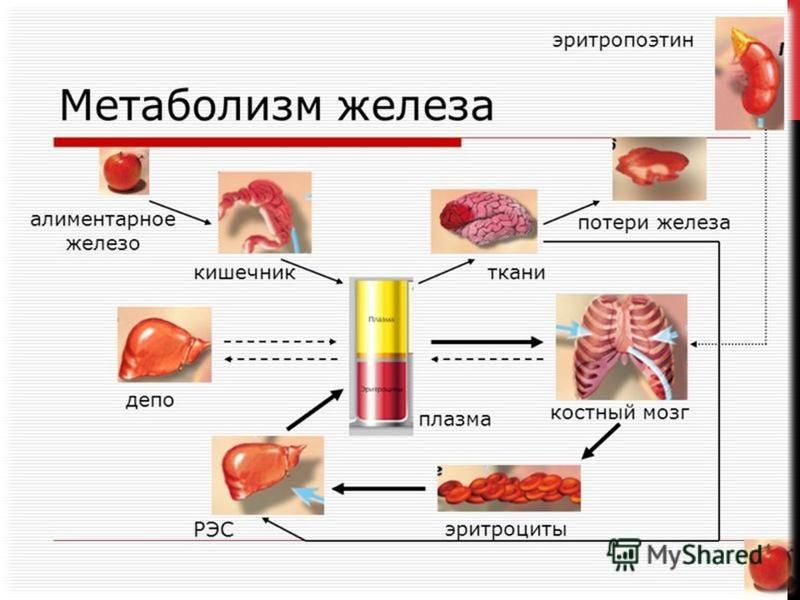
Эритропоэтин физиологический стимулятор эритропоэза. Секретируется в почках и в пери синусоидальных клетках печени. Производство эритропоэтина печенью преобладает в эмбриональный и перинатальный периоды, в то время как почечная секреция преобладает в течение зрелого возраста. Он активирует митоз и созревание эритроцитов из клеток- предшественников эритроцитарного ряда.

3
Секреция эритропоэтина почками усиливается при кровопотере, различных анемических состояниях (железо-, фолат- и B12-дефицитных анемиях, анемиях, связанных с поражениями костного мозга и др.), при ишемии почек (например, при травматическом шоке), при гипоксических состояниях
, при ишемии почек (например, при травматическом шоке), при")
5
Секреция эритропоэтина почками также усиливается под влиянием глюкокортикоидов, что служит одним из механизмов быстрого повышения уровня гемоглобина и кислород-снабжающей способности крови при стрессовых состояниях. Уровень гемоглобина и количество эритроцитов в крови повышаются уже через несколько часов после введения экзогенного эритропоэтина

6
Эритропоэтин вызывает усиленное потребление костным мозгом железа, меди, витамина B12 и филатов, которое приводит к снижению уровней железа, меди и витамина B12 в плазме крови, а также снижению уровней транспортных белков ферритина и транскобаламина Эритропоэтин повышает системное артериальное давление. Он также увеличивает вязкость крови за счёт увеличения соотношения эритроцитарной массы к плазме крови.

7
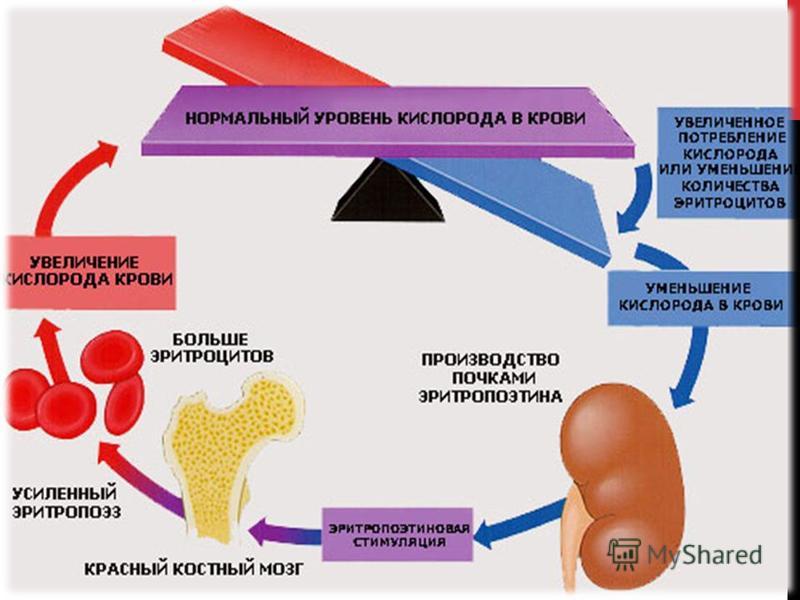
Механизм образования эритропоэтина Определяющим в образовании эритропоэтина является кислородный режим в целом организме и в частности почек. Структурной основой для выполнения этой функции является гем содержащий белок цитохром. Оксиформа этого белка тормозит выработку ИГФ-1 (индуцируемый гипоксией фактор), что происходит при снижении давления в почках от 40 до 20 мм рт. ст. Восстановленная форма приводит к нарастанию активности ИГФ-1, вследствие чего развивается экспрессия эритропоэтина. Через активацию ферментов (фосфолипазы, которая увеличивает активность простагландинов) происходит стимуляция выработки эритропоэтина.

8
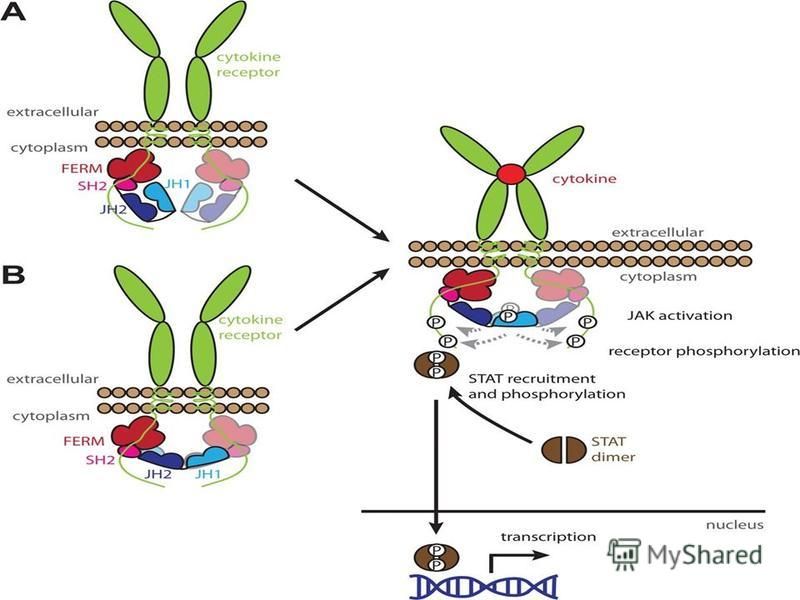
Было показано, что эритропоэтин оказывает свои эффекты путём связывания с рецептором эритропоэтина (EpoR). ЕРО связывается с рецептором эритропоэтина на поверхности клеток- предшественников и активирует JAK2 сигнальный каскад
. ЕРО связывается с рецептором эритропоэтина на поверхности клеток- предшественников и активирует JAK2 сигнальный каскад")
10
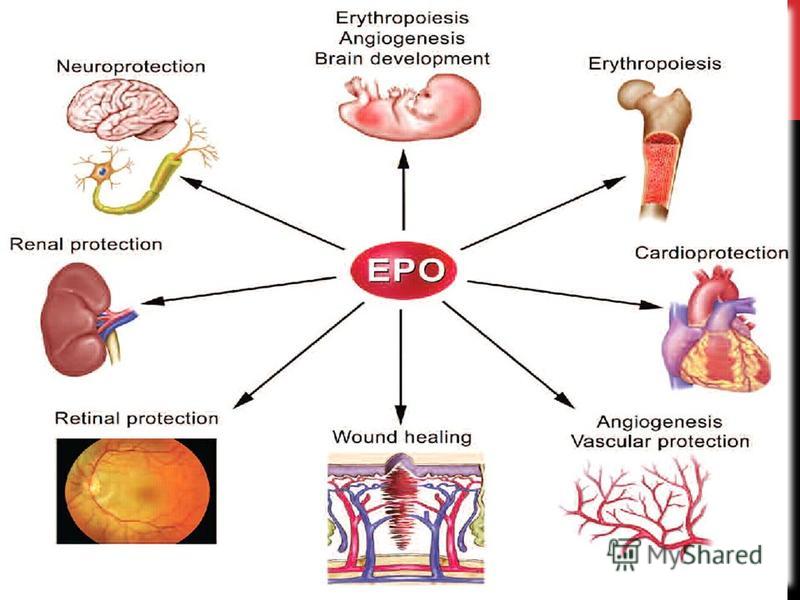
Высокоактивное проявление рецептора эритропоэтина определяет местонахождение эритроидных клеток- предшественников. Клетки-предшественники обладают высоким уровнем восприимчивости к эритропоэтину. Хотя есть сведения, что рецепторы эритропоэтина находятся в ряде других тканей (в сердце, в мышцах, в почках, в нервной ткани), достоверность результатов этих исследований искажается за счет наличия антител (anti-EpoR). Эксперименты, проводимые в контролируемых условиях, не подтвердили наличия рецептора в этих тканях. В крови, эритроциты сами не реагируют на рецептор эритропоэтина. Тем не менее, была найдена косвенная зависимость продолжительности жизни эритроцитов в крови от уровня эритропоэтина в плазме крови. Высокоактивное проявление рецептора эритропоэтина определяет местонахождение эритроидных клеток- предшественников. Клетки-предшественники обладают высоким уровнем восприимчивости к эритропоэтину. Хотя есть сведения, что рецепторы эритропоэтина находятся в ряде других тканей (в сердце, в мышцах, в почках, в нервной ткани), достоверность результатов этих исследований искажается за счет наличия антител (anti-EpoR). Эксперименты, проводимые в контролируемых условиях, не подтвердили наличия рецептора в этих тканях. В крови, эритроциты сами не реагируют на рецептор эритропоэтина. Тем не менее, была найдена косвенная зависимость продолжительности жизни эритроцитов в крови от уровня эритропоэтина в плазме крови.

11
Эритропоэтин – цитокин гликопротеинов ой природы, продуцируется интерстициальными клетками юкстагломерулярного аппарата почек и макрофагальными элементами костного мозга, печени, селезенки. Действует на КОЕ-Э, БОЕ-Э и морфологически распознаваемые клетки красной крови, регулирует интенсивность их пролиферации, определяет число митозов, совершаемых в процессе дифференцировки клетками данного класса, ускоряет построение гемоглобина, способствует освобождению ретикулоцитов из костного мозга

12
Эритропоэтин подавляет апоптоз посредством индукции экспрессии гена Bcl-XL, члена семейства Bcl-2, вовлеченного в защиту от программированной клеточной гибели. Экспрессия Bcl-XL очень низка во время ранней эритроидной дифференцировки и увеличивается в средних и поздних эритроидных клетках одновременно с потерей зависимости от эритропоэтина. Таким образом, уровень эритропоэтина детерминирует судьбу клеток эритроидной дифференцировки путем поддержания выживания ранних эритроидных клеток. Основной стимул для выработки эритропоэтина – гипоксия. Продукция эритроцитов в костном мозге под влиянием эритропоэтина может возрасти в 4-5 раз за 9-14 суток.

14
1. Ускорение и усиление перехода стволовых клеток в эритробласты; 2. Увеличение числа митозов клеток эритроидного ряда; 3. Исключение одного или нескольких циклов митотических делений; 4. Ускорение созревания неделящихся клеток - нормобластов, ретикулоцитов 5. Эритропоэтин продлевает срок жизни незрелых предшественников клеток эритроидного ряда МЕХАНИЗМ ДЕЙСТВИЯ ЭРИТРОПОЭТИНА

16
Подтверждена унитарная теория, созданная крупным русским ученым А.А Максимовым. Доказано, что для всех ростков крови существует одна родоначальная клетка. В схеме кроветворения, предложенной И.Л. Чертковым и А.И. Воробьевым, родоначальная клетка, дающая начало всем росткам кроветворной системы, соответствует I классу.

17
Эмбриональный гемопоэз делится на три периода в зависимости от времени и места протекания. Это периоды в определенной степени перекрываются: мегалобластический (внезародышевый) период й месяцы эмбриогенеза; гепато-тима-либеральный период й месяцы эмбриогенеза; медулло-тима-лимфатический период – 5-10-й месяцы эмбриогенеза.
 период - 1-2-й месяцы эмбриогенеза; гепато-тима-либеральный период - 2-5-")
18
Мегалобластический период начинается со 2-3 недели внутриутробной жизни в мезенхиме желточного мешка. В результате интенсивного деления клеток в мезенхиме образуются кровяные островки, клетки которых дифференцируются в двух направлениях: ангиобласты, лежащие по периферии, превращаются в эндотелий и образуют стенки первичных кровеносных сосудов; стволовые кроветворные клетки, которые лежат в центре островков, превращаются в первичные клетки крови – бласты.

19
Большая часть бластов делится и превращается в первичные эритробласты больших размеров – мегалобласты. Мегалобласты активно делятся и начинают синтезировать и накапливать эмбриональные гемоглобины. Из оксифильных мегалобластов образуются эритроциты больших размеров – мегалоциты. Часть мегалоцитов содержат ядро, часть – является безъядерными. Процесс образования мегалоцитов называется мегалобластическим эритропоэзом. Кроме мегалоцитов в желточном мешке образуется некоторое количество безъядерных эритроцитов обычного размера - нормобластический эритропоэз. Образование эритроцитов в желточном мешке идёт внутри кровеносных сосудов – интраваскулярной.

20
Одновременно с эритропоэзом в желточном мешке экстраваскулярной – вне просвета сосудов - идёт гранулоцитопоэз – образуются нейтрофильные и эозинофильные гранулоциты. После образования кровеносных сосудов в теле зародыша и соединения их с сосудами желточного мешка эти клетки попадают в другие органы, участвующие в эмбриональном гемопоэзе. В дальнейшем желточный мешок постепенно редуцируется, и к 12- й неделе эмбриогенеза кроветворение в нём полностью прекращается

21
В печени кроветворение начинается на 5-6 неделях развития. Здесь образуются эритроциты, гранулоциты и тромбоциты. К концу 5-го месяца интенсивность гемопоэза в печени уменьшается, но в небольшой степени продолжается ещё несколько недель после рождения. Гемопоэз в селезёнке наиболее выражен с 4-го по 8- й месяцы внутриутробного развития. Начиная с 5-го месяца красный костный мозг постепенно становится универсальным органом кроветворения, и происходит разделение на миелопоэз и лимфопоэз

22
Постэмбриональный гемопоэз – процесс образования форменных элементов крови в ходе физиологической и репаративной регенерации после рождения. Обновление различных клеточных популяций крови необходимо, поскольку абсолютное большинство форменных элементов крови имеет короткий жизненный цикл (скорость распада эритроцитов, например, составляет 10 млн в секунду). Гемопоэз обеспечивает поддержание постоянного количества форменных элементов в периферической крови. Постэмбриональный гемопоэз протекает в миелоидной (красный костный мозг) и лимфоидных (тимус, селезенка, лимфоузлы, миндалины, аппендикс, лимфатические фолликулы) тканях.

23
I полипотентные клетки-предшественники (стволовые клетки); II частично детерминированные клетки-предшественники лимфопоэза и клетки-предшественники миелопоэза; III унипотентные клетки-предшественники; IV морфологически распознаваемые пролиферирующие клетки, имеющие определенные морфологические признаки; V созревающие клетки; VI зрелые клетки: эритроциты, гранулоциты (нейтрофилы, эозинофилы, базофилы, моноциты), тромбоциты.
; II частично детерминированные клетки-предшественники лимфопоэза и клетки-предшественники миелопоэза; III унипотентные клетки-предшественники; IV морфологически распознаваемые пролиферирующие")
Еще похожие презентации в нашем архиве:




")
 – процесс образования крови. Выделяют эмбриональный и постэмбриональный гемопоэз. Эмбриональный гемопоэз.")
, лейкоцитов (лейкопоэз) и тромбоцитов (тромбоцитопоэз).")

: Н 2 О – 90-92%, белки – 7%, жиры – 0,8%, глюкоза – 0,1%, минеральные.")











