Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Boris Yu. Gumilevsky. chef immunology and allergology chair of Volgograd state medical university The Experimental models in Immunology

2
In vivo systems, which involve the whole animal, provide the most natural experimental conditions. In vivo systems, which involve the whole animal, provide the most natural experimental conditions. However, in vivo systems have a myriad of unknown and uncontrollable cellular interactions that add ambiguity to the interpretation of data However, in vivo systems have a myriad of unknown and uncontrollable cellular interactions that add ambiguity to the interpretation of data

3
in vitro systems, in which defined populations of lymphocytes are studied under controlled and consequently repeatable conditions; in vitro systems, in which defined populations of lymphocytes are studied under controlled and consequently repeatable conditions; in vitro systems can be simplified to the extent that individual cellular interactions can be studied effectively. Yet they have their own limitations, the most notable of which is their artificiality. in vitro systems can be simplified to the extent that individual cellular interactions can be studied effectively. Yet they have their own limitations, the most notable of which is their artificiality.

4
The study of the immune system in vertebrates requires suitable animal models. The choice of an animal depends on its suitability for attaining a particular research goal. The study of the immune system in vertebrates requires suitable animal models. The choice of an animal depends on its suitability for attaining a particular research goal. If large amounts of antiserum are sought, a rabbit, goat, sheep, or horse might be an appropriate experimental animal. If large amounts of antiserum are sought, a rabbit, goat, sheep, or horse might be an appropriate experimental animal. If the goal is development of a protective vaccine, the animal chosen must be susceptible to the infectious agent so that the efficacy of the vaccine can be assessed. If the goal is development of a protective vaccine, the animal chosen must be susceptible to the infectious agent so that the efficacy of the vaccine can be assessed.

5
Mice or rabbits can be used for vaccine development if they are susceptible to the pathogen. But if growth of the infectious agent is limited to humans and primates, vaccine development may require the use of monkeys, chimpanzees, or baboons. Mice or rabbits can be used for vaccine development if they are susceptible to the pathogen. But if growth of the infectious agent is limited to humans and primates, vaccine development may require the use of monkeys, chimpanzees, or baboons. For most basic research in immunology, mice have been the experimental animal of choice. They are easy to handle, are genetically well characterized, and have a rapid breeding cycle. The immune system of the mouse has been characterized more extensively than that of any other species For most basic research in immunology, mice have been the experimental animal of choice. They are easy to handle, are genetically well characterized, and have a rapid breeding cycle. The immune system of the mouse has been characterized more extensively than that of any other species

6
Inbred Strains Can Reduce Experimental Variation To control experimental variation caused by differences in the genetic backgrounds of experimental animals, immunologists often work with inbred strains-that is, genetically identical animals produced by inbreeding. To control experimental variation caused by differences in the genetic backgrounds of experimental animals, immunologists often work with inbred strains-that is, genetically identical animals produced by inbreeding.

7
The rapid breeding cycle of mice makes them particularly well suited for the production of inbred strains, which are developed by repeated inbreeding between brother and sister littermates. The rapid breeding cycle of mice makes them particularly well suited for the production of inbred strains, which are developed by repeated inbreeding between brother and sister littermates. In this way the heterozygosity of alleles that is normally found in randomly outbreed mice is replaced by homozygosity at all loci. In this way the heterozygosity of alleles that is normally found in randomly outbreed mice is replaced by homozygosity at all loci. Repeated inbreeding for 20 generations usually yields an inbred strain whose progeny are homozygous at more than 98% of all loci. Repeated inbreeding for 20 generations usually yields an inbred strain whose progeny are homozygous at more than 98% of all loci.

8
Because inbred strains of animals are genetically identical (syngeneic) within that strain, their immune responses can be studied in the absence of variables introduced by individual genetic differences-an invaluable property. Because inbred strains of animals are genetically identical (syngeneic) within that strain, their immune responses can be studied in the absence of variables introduced by individual genetic differences-an invaluable property.
 within that strain, their immune responses can be studied in the absence of variables introduced by individual genetic differences-an invaluable property. Because inbred strains")
9
With inbred strains, lymphocyte subpopulations isolated from one animal can be injected into another animal of the same strain without eliciting a rejection reaction. With inbred strains, lymphocyte subpopulations isolated from one animal can be injected into another animal of the same strain without eliciting a rejection reaction. This type of experimental system permitted immunologists to first demonstrate that lymphocytes from an antigen-primed animal could transfer immunity to an unprimed syngeneic recipient. This type of experimental system permitted immunologists to first demonstrate that lymphocytes from an antigen-primed animal could transfer immunity to an unprimed syngeneic recipient.

12
Adoptive-Transfer Systems Permit the in Vivo Examination of Isolated Cell Populations In some experiments, it is important to eliminate the immune responsiveness of the syngeneic host so that the response of only the transferred lymphocytes can be studied in isolation. In some experiments, it is important to eliminate the immune responsiveness of the syngeneic host so that the response of only the transferred lymphocytes can be studied in isolation.

13
This can be accomplished by a technique called adoptive transfer: first, the syngeneic host is exposed to x-rays that kill its lymphocytes; then the donor immune cells are introduced. This can be accomplished by a technique called adoptive transfer: first, the syngeneic host is exposed to x-rays that kill its lymphocytes; then the donor immune cells are introduced. Subjecting a mouse to high doses of x-rays ( rads) can kill 99.99% of its lymphocytes, after which the activities of lymphocytes transplanted from the spleen of a syngeneic donor can be studied without interference from host lymphocytes. Subjecting a mouse to high doses of x-rays ( rads) can kill 99.99% of its lymphocytes, after which the activities of lymphocytes transplanted from the spleen of a syngeneic donor can be studied without interference from host lymphocytes.

14
If the host's hematopoietic cells might influence an adoptive-transfer experiment, then higher x-ray levels ( rads) are used to eliminate the entire haematopoietic system. If the host's hematopoietic cells might influence an adoptive-transfer experiment, then higher x-ray levels ( rads) are used to eliminate the entire haematopoietic system. Mice irradiated with such doses will die unless reconstituted with bone marrow from a syngeneic donor. Mice irradiated with such doses will die unless reconstituted with bone marrow from a syngeneic donor.
 are used to eliminate the entire haematopoietic system. If the host's hematopoietic cells might influence an adoptive-transfer")
15
The adoptive-transfer system has enabled immunologists to study the development of injected lymphoid stem cells in various organs of the recipient, and have facilitated the study of various populations of lymphocytes and of the cellular interactions required to generate an immune response.

16
Such experiments, for instance, first enabled immunologists to show that a T helper cell is necessary for B-cell activation in the humoral response. Such experiments, for instance, first enabled immunologists to show that a T helper cell is necessary for B-cell activation in the humoral response. In these experiments, adoptive transfer of purified B cells or purified T cells did not produce antibody in the irradiated host. Only when both cell populations were transferred was antibody produced in response to antigen. In these experiments, adoptive transfer of purified B cells or purified T cells did not produce antibody in the irradiated host. Only when both cell populations were transferred was antibody produced in response to antigen.

17
SCID Mice and SCID-Human Mice Are a Valuable Animal Model for immunodeficiency An autosomal recessive mutation resulting in severe combined immunodeficiency disease (SCID) developed spontaneously in a strain of mice called CB-17. These mice fail to develop mature T and B cells and consequently are severely compromised immunologically. The absence of functional T and B cells enables these mice to accept foreign cells and grafts from other strains of mice or even from other species. An autosomal recessive mutation resulting in severe combined immunodeficiency disease (SCID) developed spontaneously in a strain of mice called CB-17. These mice fail to develop mature T and B cells and consequently are severely compromised immunologically. The absence of functional T and B cells enables these mice to accept foreign cells and grafts from other strains of mice or even from other species.
 developed spontaneously in a strain of mice called CB-17. These mice fail to d")
18
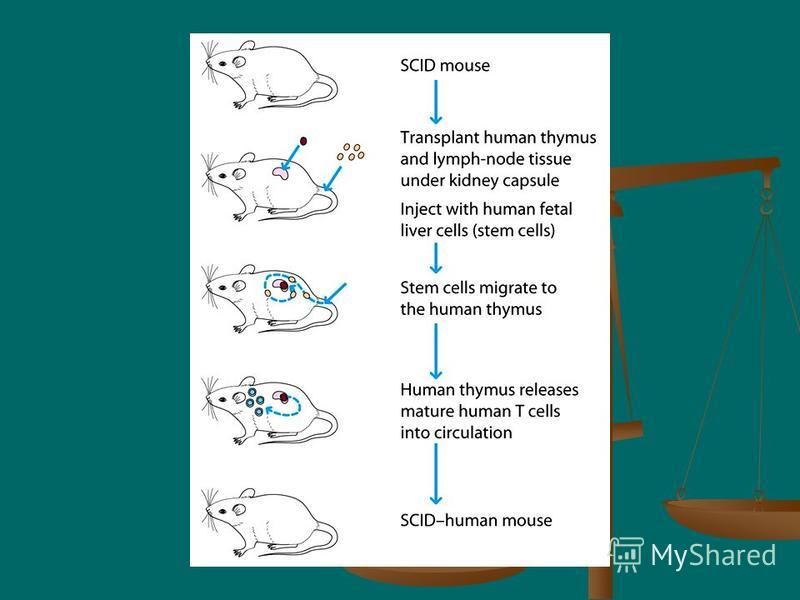
When normal bone marrow cells are injected into SCID mice, normal T and B cells develop, and the mice are cured of their immunodeficiency. When normal bone marrow cells are injected into SCID mice, normal T and B cells develop, and the mice are cured of their immunodeficiency. This finding has made SCID mice a valuable model system for the study of immunodeficiency and the process of differentiation of bone-marrow stem cells into mature T or B cells. This finding has made SCID mice a valuable model system for the study of immunodeficiency and the process of differentiation of bone-marrow stem cells into mature T or B cells. Interest in SCID mice mushroomed when it was found that they could be used to study the human immune system. Interest in SCID mice mushroomed when it was found that they could be used to study the human immune system.

20
The implanted human fetal liver contains immature lymphocytes (stem cells), which migrate to the implanted human tissues, where they mature into T and B cells, producing a SCIDhuman mouse. The implanted human fetal liver contains immature lymphocytes (stem cells), which migrate to the implanted human tissues, where they mature into T and B cells, producing a SCIDhuman mouse. Because the human lymphocytes are exposed to mouse antigens while they are still immature, they later recognize mouse cells as self and do not mount an immunologic response against the mouse host. Because the human lymphocytes are exposed to mouse antigens while they are still immature, they later recognize mouse cells as self and do not mount an immunologic response against the mouse host.
, which migrate to the implanted human tissues, where they mature into T and B cells, producing a SCIDhuman mouse. The implanted human fetal liver contains immature lymphocytes")
21
The beauty of the SCID-human mouse is that it enables one to study human lymphocytes within an animal model. The beauty of the SCID-human mouse is that it enables one to study human lymphocytes within an animal model. This valuable system has proved useful in research on the development of various lymphoid cells and also as an important animal model in AIDS research, since mouse lymphocytes cannot be infected with HIV, whereas the lymphocytes of a SCID-human mouse are readily infected. This valuable system has proved useful in research on the development of various lymphoid cells and also as an important animal model in AIDS research, since mouse lymphocytes cannot be infected with HIV, whereas the lymphocytes of a SCID-human mouse are readily infected.

22
Cell-Culture Systems The complexity of the cellular interactions that generate an immune response has led immunologists to rely heavily on various types of in vitro cell-culture systems. A variety of cells can be cultured, including primary lymphoid cells, cloned lymphoid cell lines, and hybrid cells. The complexity of the cellular interactions that generate an immune response has led immunologists to rely heavily on various types of in vitro cell-culture systems. A variety of cells can be cultured, including primary lymphoid cells, cloned lymphoid cell lines, and hybrid cells.

23
Primary Lymphoid Cell Cultures Primary lymphoid cell cultures can be obtained by isolating lymphocytes directly from blood or lymph or from various lymphoid organs by tissue dispersion. The lymphocytes can then be grown in a chemically defined basal medium (containing saline, sugars, amino acids, vitamins, trace elements, and other nutrients) to which various serum supplements are added. Primary lymphoid cell cultures can be obtained by isolating lymphocytes directly from blood or lymph or from various lymphoid organs by tissue dispersion. The lymphocytes can then be grown in a chemically defined basal medium (containing saline, sugars, amino acids, vitamins, trace elements, and other nutrients) to which various serum supplements are added.

24
Because in vitro culture techniques require from 10- to 100-fold fewer lymphocytes than do typical in vivo techniques, they have enabled immunologists to assess the functional properties of minor subpopulations of lymphocytes. Because in vitro culture techniques require from 10- to 100-fold fewer lymphocytes than do typical in vivo techniques, they have enabled immunologists to assess the functional properties of minor subpopulations of lymphocytes.

25
It was by means of cell-culture techniques, for example, that immunologists were first able to define the functional differences between CD4+ T helper cells and CD8+ T cytotoxic cells. Cell-culture techniques have also been used to identify numerous cytokines involved in the activation, growth, and differentiation of various cells involved in the immune response.

26
Cloned Lymphoid Cell Lines A primary lymphoid cell culture comprises a heterogeneous group of cells that can be propagated only for a limited time. This heterogeneity can complicate the interpretation of experimental results. To avoid these problems, immunologists use cloned lymphoid cell lines and hybrid cells. A primary lymphoid cell culture comprises a heterogeneous group of cells that can be propagated only for a limited time. This heterogeneity can complicate the interpretation of experimental results. To avoid these problems, immunologists use cloned lymphoid cell lines and hybrid cells.

27
Normal mammalian cells generally have a finite life span in culture; that is, after a number of population doublings characteristic of the species and cell type, the cells stop dividing. In contrast, tumor cells or normal cells that have undergone transformation induced by chemical carcinogens or viruses can be propagated indefinitely in tissue culture; thus, they are said to be immortal. Such cells are referred to as cell lines.

28
The first cell line-the mouse fibroblast L cell- was derived in the 1940s from cultured mouse subcutaneous connective tissue by exposing the cultured cells to a chemical carcinogen, methylcholanthrene, over a 4-month period. The first cell line-the mouse fibroblast L cell- was derived in the 1940s from cultured mouse subcutaneous connective tissue by exposing the cultured cells to a chemical carcinogen, methylcholanthrene, over a 4-month period. In the 1950s, another important cell line, the HeLa cell, was derived by culturing human cervical cancer cells. In the 1950s, another important cell line, the HeLa cell, was derived by culturing human cervical cancer cells.

31
By culturing normal T lymphocytes with antigen in the presence of IL-2, clones of antigen-specific T lymphocytes could be isolated. These individual clones could be propagated and studied in culture and even frozen for storage. After thawing, the clones continued to grow and express their original antigenspecific functions. Development of cloned lymphoid cell lines has enabled immunologists to study a number of events that previously could not be examined. By culturing normal T lymphocytes with antigen in the presence of IL-2, clones of antigen-specific T lymphocytes could be isolated. These individual clones could be propagated and studied in culture and even frozen for storage. After thawing, the clones continued to grow and express their original antigenspecific functions. Development of cloned lymphoid cell lines has enabled immunologists to study a number of events that previously could not be examined.

32
The genetic changes corresponding to different maturational stages can be studied in cell lines that appear to be "frozen" at different stages of differentiation. Cell lines have also been useful in studying the soluble factors produced by lymphoid cells. Some cell lines secrete large quantities of various cytokines; other lines express membrane receptors for particular cytokines. These cell lines have been used by immunologists to purify various cytokines and their receptors and eventually to clone their genes. The genetic changes corresponding to different maturational stages can be studied in cell lines that appear to be "frozen" at different stages of differentiation. Cell lines have also been useful in studying the soluble factors produced by lymphoid cells. Some cell lines secrete large quantities of various cytokines; other lines express membrane receptors for particular cytokines. These cell lines have been used by immunologists to purify various cytokines and their receptors and eventually to clone their genes.

33
Hybrid Lymphoid Cell Lines In somatic-cell hybridization, immunologists fuse normal B or T lymphocytes with tumor cells, obtaining hybrid cells, or heterokaryons, containing nuclei from both parent cells. Random loss of some chromosomes and subsequent cell proliferation yield a clone of cells that contain a single nucleus with chromosomes from each of the fused cells; such a clone is called a hybridoma. In somatic-cell hybridization, immunologists fuse normal B or T lymphocytes with tumor cells, obtaining hybrid cells, or heterokaryons, containing nuclei from both parent cells. Random loss of some chromosomes and subsequent cell proliferation yield a clone of cells that contain a single nucleus with chromosomes from each of the fused cells; such a clone is called a hybridoma.

35
B-cell hybridomas that secrete antibody with a single antigenic specificity, called monoclonal antibody, in reference to its derivation from a single clone, have revolutionized not only immunology but biomedical research as well as the clinical laboratory. T-cell hybridomas can also be obtained by fusing T lymphocytes with cancerous T-cell lymphomas.

36
Cloned Genes Tranferred into Mouse Embryos Allow in Vivo Analysis of Gene Function Development of techniques to introduce cloned foreign genes (called transgenes) into mouse embryos has permitted immunologists to study the effects of immune-system genes in vivo. Development of techniques to introduce cloned foreign genes (called transgenes) into mouse embryos has permitted immunologists to study the effects of immune-system genes in vivo. If the introduced gene integrates stably into the germ-line cells, it will be transmitted to the offspring. If the introduced gene integrates stably into the germ-line cells, it will be transmitted to the offspring.
 into mouse embryos has permitted immunologists to study the effects of immune-system ge")
38
With transgenic mice, immunologists have been able to study the expression of a given gene in a living animal. Although all the cells in a transgenic animal contain the transgene, differences in the expression of the transgene in different tissues has shed light on mechanisms of tissue-specific gene expression. With transgenic mice, immunologists have been able to study the expression of a given gene in a living animal. Although all the cells in a transgenic animal contain the transgene, differences in the expression of the transgene in different tissues has shed light on mechanisms of tissue-specific gene expression.

39
Because a transgene is integrated into the chromosomal DNA within the one- celled mouse embryo, it will be integrated into both somatic cells and germ-line cells. The resulting transgenic mice thus can transmit the transgene to their offspring as a Mendelian trait. In this way, it has been possible to produce lines of transgenic mice in which every member of a line contains the same transgene. Because a transgene is integrated into the chromosomal DNA within the one- celled mouse embryo, it will be integrated into both somatic cells and germ-line cells. The resulting transgenic mice thus can transmit the transgene to their offspring as a Mendelian trait. In this way, it has been possible to produce lines of transgenic mice in which every member of a line contains the same transgene.

40
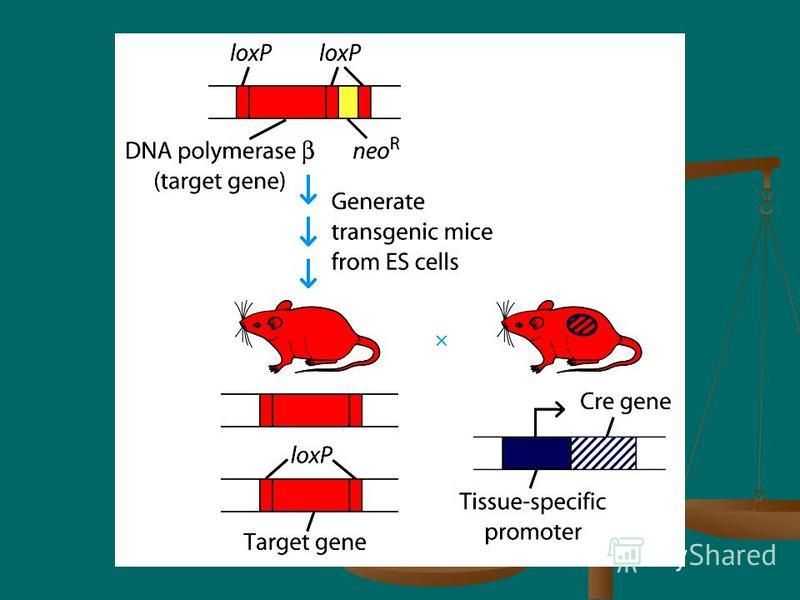
One of the limitations with transgenic mice is that the transgene is integrated randomly within the genome. One of the limitations with transgenic mice is that the transgene is integrated randomly within the genome. To circumvent this limitation, researchers have developed a technique in which a desired gene is targeted to specific sites within the germ line of a mouse. The primary use of this technique has been to replace a normal gene with a mutant allele or a disrupted form of the gene, thus knocking out the gene's function. Transgenic mice that carry such a disrupted gene, called knockout mice To circumvent this limitation, researchers have developed a technique in which a desired gene is targeted to specific sites within the germ line of a mouse. The primary use of this technique has been to replace a normal gene with a mutant allele or a disrupted form of the gene, thus knocking out the gene's function. Transgenic mice that carry such a disrupted gene, called knockout mice

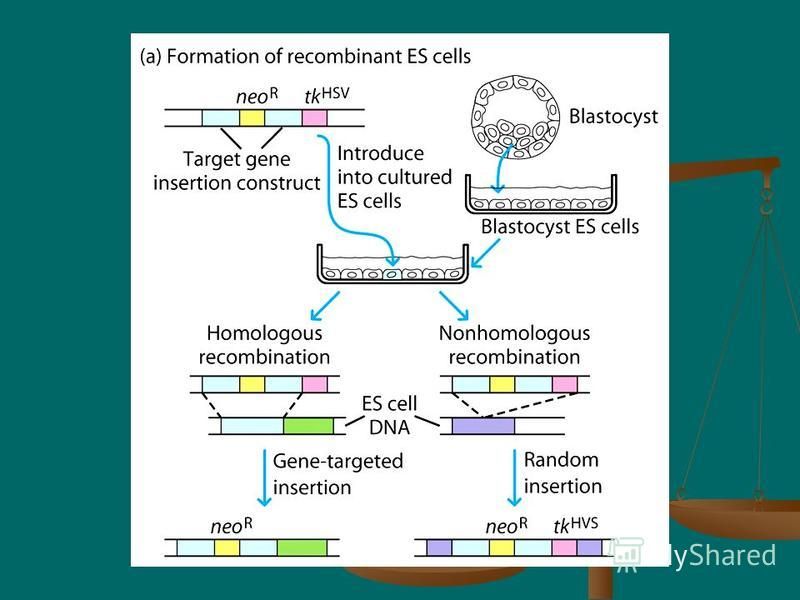
41
Production of gene-targeted knockout mice involves the following steps: Isolation and culturing of embryonic stem (ES) cells from the inner cell mass of a mouse blastocyst Isolation and culturing of embryonic stem (ES) cells from the inner cell mass of a mouse blastocyst Introduction of a mutant or disrupted gene into the cultured ES cells and selection of homologous recombinant cells in which the gene of interest has been knocked out (i.e., replaced by a nonfunctional form of the gene) Introduction of a mutant or disrupted gene into the cultured ES cells and selection of homologous recombinant cells in which the gene of interest has been knocked out (i.e., replaced by a nonfunctional form of the gene)
 cells from the inner cell mass of a mouse blastocyst Isolation and culturing of embryonic stem (ES) cells from the inner cell mass")
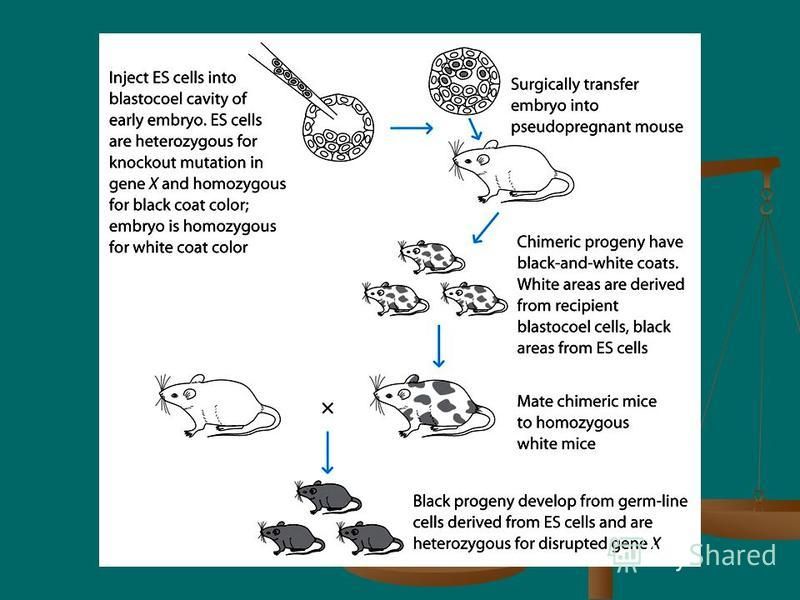
42
Injection of homologous recombinant ES cells into a recipient mouse blastocyst and surgical implantation of the blastocyst into a pseudo-pregnant mouse Injection of homologous recombinant ES cells into a recipient mouse blastocyst and surgical implantation of the blastocyst into a pseudo-pregnant mouse Mating of chimeric offspring heterozygous for the disrupted gene to produce homozygous knockout mice. Mating of chimeric offspring heterozygous for the disrupted gene to produce homozygous knockout mice.

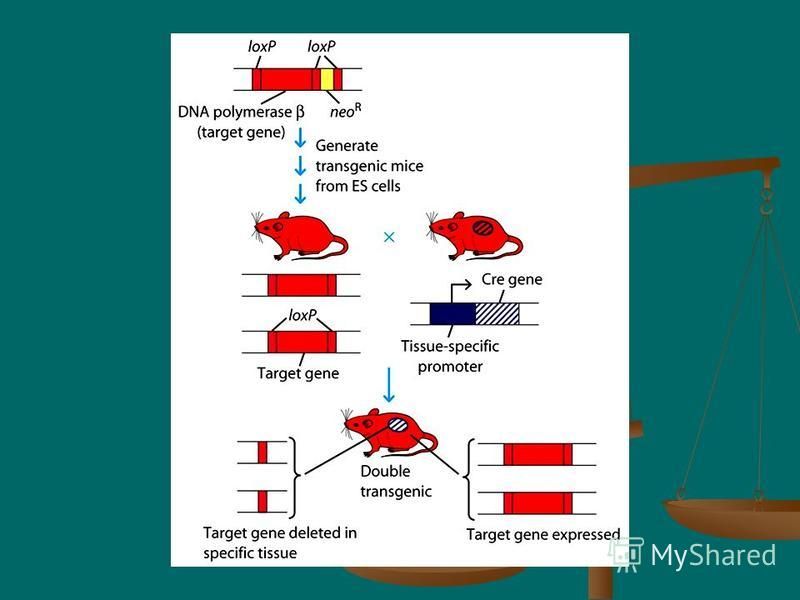
46
"Knock-In" Technology Allows the Replacement of an Endogenous Gene In addition to deleting a gene of choice, it also is possible to replace the endogenous gene with a mutated form of that gene. "Knock-In" Technology Allows the Replacement of an Endogenous Gene In addition to deleting a gene of choice, it also is possible to replace the endogenous gene with a mutated form of that gene. As in the strategy for knocking out a gene, DNA constructs that carry mutations in a particular gene can be exchanged for the endogenous gene. It also is possible to replace all of an endogenous gene with a DNA sequence of choice. As in the strategy for knocking out a gene, DNA constructs that carry mutations in a particular gene can be exchanged for the endogenous gene. It also is possible to replace all of an endogenous gene with a DNA sequence of choice.

47
In a recent report, for example, the CD4 gene was replaced with the one for alfa-galactosidase. In these experiments, the CD4 promoter was left intact to drive the expression of alfa-galactosidase, which catalyzes the color change of certain reporter chemicals to blue. In a recent report, for example, the CD4 gene was replaced with the one for alfa-galactosidase. In these experiments, the CD4 promoter was left intact to drive the expression of alfa-galactosidase, which catalyzes the color change of certain reporter chemicals to blue.

48
Because the CD4 promoter drove the expression of alfa-galactosidase, only those thymic cells destined to express CD4 turned blue in the presence of the reporter chemicals. Data from these experiments were useful in tracing CD4/CD8 lineage commitment in developing T cells.

49
The large thank for your attention!

Еще похожие презентации в нашем архиве:









 Transplantation Clones Cloning is the moving of an organ from one body to another or from a donor site on the patients.")

![Genetics Genetics (from Ancient Greek γενετικός genetikos, genitive and that from γένεσις genesis, origin),[1][2][3] a discipline of biology, is the.](/thumbs/17/1180906/big_thumb.jpg "Genetics Genetics (from Ancient Greek γενετικός genetikos, genitive and that from γένεσις genesis, origin),[1][2][3] a discipline of biology, is the.")



, I (for infectious) and R (for recovered). Therefore, this.")



 The virus compromises the bodys ability to handle disease and causes AIDS. AIDS~ (Acquired Immune Deficiency.")


 is recognized as one of the key competencies necessary for success in.")






