Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Биологическое окисление

2
Переход энергии

3
Окисление и восстановление являются химическими реакциями, описывающими, соответственно, потерю и приобретение атомами электронов, что происходит сопряжено Окисление и восстановление являются химическими реакциями, описывающими, соответственно, потерю и приобретение атомами электронов, что происходит сопряжено

4
Биологическое окисление – окислительно-восстановительные реакции, протекающие в биологических системах при участии ферментов Биологическое окисление – окислительно-восстановительные реакции, протекающие в биологических системах при участии ферментов Чаще всего акцептором электронов является кислород, донором - водород Чаще всего акцептором электронов является кислород, донором - водород

5
Переход электронов от восстановителя к окислителю

6
Количество водорода, входящего в состав белков, жиров и углеводов будет рассматриваться как количество потенциальной энергии вещества Количество водорода, входящего в состав белков, жиров и углеводов будет рассматриваться как количество потенциальной энергии вещества Энергия выделяется в реакциях катаболизма!!! Энергия выделяется в реакциях катаболизма!!!

7
Общая схема катаболизма

8
Первый этап катаболизма (подготовительный)
")
9
Биологическая роль подготовительного этапа катаболизма - распад полимеров (белков, жиров, углеводов) в ЖКТ с помощью пищеварительных ферментов до мономеров Биологическая роль подготовительного этапа катаболизма - распад полимеров (белков, жиров, углеводов) в ЖКТ с помощью пищеварительных ферментов до мономеров
 в ЖКТ с помощью пищеварительных ферментов до мономеров Биологическая роль подготовительного этапа катаболизма - распад полимеров (белков, жиров, угле")
10
Второй этап катаболизма (внутриклеточный)
")
11
Биологическая роль внутриклеточного этапа катаболизма – поступление мономеров в клетку (аминокислот, жирных кислот, глюкозы) и превращение их в центральный метаболит – Биологическая роль внутриклеточного этапа катаболизма – поступление мономеров в клетку (аминокислот, жирных кислот, глюкозы) и превращение их в центральный метаболит – Ацетил –Ко-А
 и превращение их в центральный метаболит – Биологическая роль внутриклеточного этапа катаболизма – поступление мономеров в кл")
12
Третий этап катаболизма (митохондриальный)
")
13
Биологическая роль митохондриального этапа катаболизма – получение энергии Биологическая роль митохондриального этапа катаболизма – получение энергии Ацетил-Ко-А вступает в цикл трикарбоновых кислот (ЦТК), в результате чего восстанавливаются коферменты НАДН и ФАДН 2, следующие в дыхательную цепь Ацетил-Ко-А вступает в цикл трикарбоновых кислот (ЦТК), в результате чего восстанавливаются коферменты НАДН и ФАДН 2, следующие в дыхательную цепь При транспорте электронов и протонов водорода в дыхательной цепи синтезируется АТФ. При транспорте электронов и протонов водорода в дыхательной цепи синтезируется АТФ.
, в результате чего восстанавливаются кофермен")
14
Цикл трикарбоновых кислот

15
Ключевые моменты ЦТК: Локализация процесса – матрикс митохондрий Локализация процесса – матрикс митохондрий Центральный метаболит ацетил-КоА взаимодействует с оксалоацетатом до цитрата, который опять превращается в оксалоацетат, образуя цикл Центральный метаболит ацетил-КоА взаимодействует с оксалоацетатом до цитрата, который опять превращается в оксалоацетат, образуя цикл Проходит серия дегидрогеназных реакций, из которых три НАД-зависимых, одна ФАД-зависимая Проходит серия дегидрогеназных реакций, из которых три НАД-зависимых, одна ФАД-зависимая Осуществляется одна реакция субстратного фосфорилирования, где энергия выделяется при распаде молекулы субстрата без участия дыхательной цепи Осуществляется одна реакция субстратного фосфорилирования, где энергия выделяется при распаде молекулы субстрата без участия дыхательной цепи Выделяют два регулируемых фермента: цитратсинтаза, изоцитратдегидрогеназа Выделяют два регулируемых фермента: цитратсинтаза, изоцитратдегидрогеназа

16
Регуляция цикла ЦТК Два регулируемых фермента: цитратсинтаза, изоцитратдегидрогеназа Два регулируемых фермента: цитратсинтаза, изоцитратдегидрогеназа Оба фермента аллостерическийййй ингибируются избытком АТФ и НАДН 2 Оба фермента аллостерическийййй ингибируются избытком АТФ и НАДН 2 Оба фермента аллостерическийййй активируются АДФ и НАД + Оба фермента аллостерическийййй активируются АДФ и НАД +

17
Биологическая роль ЦТК: ЦТК – поставщик НАДН и ФАДН 2 в дыхательную цепь, в связи с этим главная роль – ЭНЕРГЕТИЧЕСКАЯ ЦТК – универсальный терминальный этап катаболизма веществ всех классов ЦТК – играет важную роль в процессах анаболизма: из цитрата => синтез жирных кислот, из -кетоглутарата и оксалоацетата => синтез аминокислот, из оксалоацетата => синтез углеводов, из сукцинил-КоА => синтез гема

18
Виды биологического окисления Двуэлектронное окисление. Двуэлектронное окисление. При сопряженном окислении-восстановлении передаются 2 электрона. Биологическая роль– энергетическая. Чаще всего эти реакции катализируются ферментами дегидрогеназами

19
Одноэлектронное окисление. Одноэлектронное окисление. При сопряженном окислении-восстановлении передаются 1 электрон. Биологическая роль– сигнальная. Чаще всего эти реакции катализируются ферментами оксидазами.

21
Двуэлектронное окисление (дыхательная цепь, электротранспортная цепь, тканевое дыхание)
")
22
Участники окислительно- восстановительных реакций (дыхательной цепи) НАД + - НАДН 2 НАД + - НАДН 2 ФАД - ФАДН 2 ФАД - ФАДН 2 Убихинон (Q) – убихинон восстановленный (QН 2 ) Убихинон (Q) – убихинон восстановленный (QН 2 ) Цитохром В Цитохром В Цитохром С Цитохром С Цитохром А и А 3 Цитохром А и А 3
 НАД + - НАДН 2 НАД + - НАДН 2 ФАД - ФАДН 2 ФАД - ФАДН 2 Убихинон (Q) – убихинон восстановленный (QН 2 ) Убихинон (Q) – убихинон восстановленный (QН 2 ) Цитохром В Цитохром В Цитохро")
23
Принцип построения дыхательной цепи Участники окислительно-восстановительных реакций (дыхательной цепи), являющиеся переносчиками электронов, располагаются соответственно величине окислительно- восстановительного потенциала Участники окислительно-восстановительных реакций (дыхательной цепи), являющиеся переносчиками электронов, располагаются соответственно величине окислительно- восстановительного потенциала
, являющиеся переносчиками электронов, располагаются соответственно величине окислительно- восстановительного потенциала Участники окислительно-вос")
24
Строение молекулы НАД (НАДН)
")
25
Строение молекулы ФАД (ФАДН 2 )
")
26
Строение убихинона (Q)
")
27
Строение цитохромов

28
Атом железа в геме может менять валентность, присоединяя или отдавая электроны: Атом железа в геме может менять валентность, присоединяя или отдавая электроны:

29
Строение АТФ

30
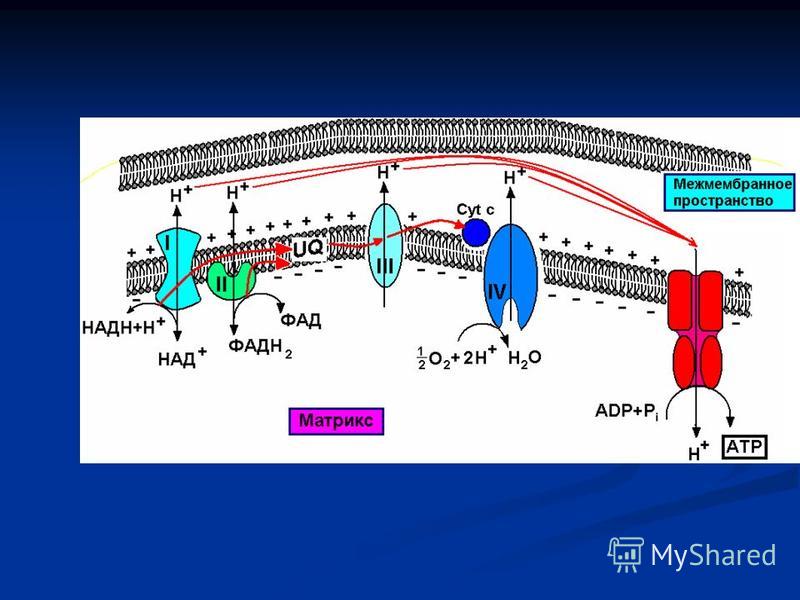
Дыхательная цепь

31
При переходе электронов от одного атома к другому, происходит выделение энергии. Выделившаяся энергия, подчиняясь I закону термодинамики, о превращении энергии из одной формы в другую, либо расходуется в виде тепла, либо аккумулируется организмом в виде макроэргических связях молекул АТФ.

32
Строение дыхательной цепи Локализация процесса – внутренняя мембрана митохондрий Локализация процесса – внутренняя мембрана митохондрий Дыхательную цепь формируют четыре комплекса, расположенные в определенной последовательности (векторно) Дыхательную цепь формируют четыре комплекса, расположенные в определенной последовательности (векторно) Принцип построения цепи - возрастание окислительно-восстановительного потенциала по мере приближения к кислороду Принцип построения цепи - возрастание окислительно-восстановительного потенциала по мере приближения к кислороду
 Дыхательну")
33
I комплекс НАДН-дегидрогеназный НАДН-дегидрогеназный кофермент – ФМН кофермент – ФМН простетическая группа – FeS простетическая группа – FeS Функция – забирает электроны от НАДН (из ЦТК) и передает ФМН, а затем убихинону - Q Функция – забирает электроны от НАДН (из ЦТК) и передает ФМН, а затем убихинону - Q
 и передает ФМН, а затем убихинону - Q Функция – забирает электр")
34
II комплекс Сукцинатдегидрогеназный Сукцинатдегидрогеназный кофермент – ФАД кофермент – ФАД простетическая группа – FeS простетическая группа – FeS Функция – забирает электроны от ФАДН 2 (из ЦТК) и передает убихинону - Q. Функция – забирает электроны от ФАДН 2 (из ЦТК) и передает убихинону - Q.
 и передает убихинону - Q. Функция – забирает электроны")
35
III комплекс Цитохромы В, С 1 Цитохромы В, С 1 кофермент – гемы кофермент – гемы простетическая группа – FeS простетическая группа – FeS Функция – забирает электроны от убихинона и передает цитохрому С Функция – забирает электроны от убихинона и передает цитохрому С

36
IV комплекс Цитохромоксидазный Цитохромоксидазный кофермент – гемы кофермент – гемы кофактор – Сu 2+ кофактор – Сu 2+ Функция – забирает электроны от цитохрома С и передает кислороду Функция – забирает электроны от цитохрома С и передает кислороду

37
Хемоосмотическая теория дыхательной цепи Цепь транспорта электронов функционирует как протонная (Н+)помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство Цепь транспорта электронов функционирует как протонная (Н+)помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи
помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство Цепь транспорта электронов функционирует как пр")
38
Перенос протонов в межмембранное пространство приводит к возникновению разности концентрации Н + с двух сторон внутренней митохондриальной мембраны: более высокая снаружи и более низкая - внутри Перенос протонов в межмембранное пространство приводит к возникновению разности концентрации Н + с двух сторон внутренней митохондриальной мембраны: более высокая снаружи и более низкая - внутри В результате возникает градиент концентрации Н + и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности внутренней мембраны В результате возникает градиент концентрации Н + и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности внутренней мембраны

40
Окислительное фосфорилирование Синтез АТФ из АДФ и фосфата за счет энергии переноса электронов по дыхательной цепи называют окислительным фосфорилированием Синтез АТФ из АДФ и фосфата за счет энергии переноса электронов по дыхательной цепи называют окислительным фосфорилированием Это совокупность двух процессов: Это совокупность двух процессов: Первый процесс – экзэргонический (выделение энергии) - перенос электронов по цепи к кислороду Второй процесс – эндергонический (затрата энергии) - фосфорилирование АДФ (синтез АТФ) Второй процесс – эндергонический (затрата энергии) - фосфорилирование АДФ (синтез АТФ)

41
Регуляция дыхательной цепи или дыхательный контроль Скорость дыхания митохондрий может контролироваться концентрацией ADP Скорость дыхания митохондрий может контролироваться концентрацией ADP Окисление и фосфорилирование жестко сопряжены, поэтому энергия, необходимая клетке для совершения работы, поставляется за счет гидролиза АТР Окисление и фосфорилирование жестко сопряжены, поэтому энергия, необходимая клетке для совершения работы, поставляется за счет гидролиза АТР Концентрация ADP при этом увеличивается, в результате создаются условия для ускорения дыхания, что и ведет к восполнению запасов АТР Концентрация ADP при этом увеличивается, в результате создаются условия для ускорения дыхания, что и ведет к восполнению запасов АТР

42
Разобщение дыхания и фосфорилирования Разобщители дыхания и фосфорилирования способны выравнивать концентрацию водорода в матриксе и межмембранном пространстве Разобщители дыхания и фосфорилирования способны выравнивать концентрацию водорода в матриксе и межмембранном пространстве Уничтожается электрохимический потенциал, и синтез АТР становится невозможным, хотя окисление субстратов при этом происходит Уничтожается электрохимический потенциал, и синтез АТР становится невозможным, хотя окисление субстратов при этом происходит Энергия дыхательной цепи в этом случае полностью рассеивается в виде теплоты Энергия дыхательной цепи в этом случае полностью рассеивается в виде теплоты

43
Разобщающим действием обладают: гормон щитовидной железы – тироксин гормон щитовидной железы – тироксин жирные кислоты жирные кислоты некоторые антибиотики (валиномицин и грамицидин) некоторые антибиотики (валиномицин и грамицидин) РБ-1-термогенин (бурый жир), участвует в терморегуляции РБ-1-термогенин (бурый жир), участвует в терморегуляции
 некоторые антибиотики (валиномицин и грамицидин) РБ-1-термогенин (б")
Еще похожие презентации в нашем архиве:

 Пластический обмен (анаболизм, ассимиляция) Совокупность реакций синтеза, которые идут с затратой энергии АТФ.")






подготовительный.")

 Кислородный ( аэробный)")

 Доцент кафедры биохимии к. б. н. Лобаева Т. А.")







 (Mg 2+ ) (2.7.1.1.) 2.Глюкозофосфатизомераза (5.3.1.9.)")
