Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Посттранскрипционный контроль

2
Хотя контроль активности большинства генов осуществляется главным образом на уровне инициации транскрипции, тем не менее и позже, в ходе передачи информации от РНК к белку, такой контроль происходит на самых разных этапах. Более того, вполне вероятно, что для некоторых генов каждая стадия их экспрессии находится под контролем. Большая часть сведений о посттранскрипционной регуляции получена лишь недавно. Один из основных путей адаптации организмов к изменяющимся условиям окружающей среды регуляция экспрессии генов. Этот процесс, детально изученный для бактерий и вирусов, заключается в специфическом взаимодействии определенных белков с различными участками ДНК, расположенными рядом с сайтами инициации транскрипции. Такие взаимодействия могут характеризоваться как позитивным (положительным), так и негативным (отрицательным) влиянием на уровень транскрипции. В эукариотических клетках используются и другие механизмы регуляции транскрипции. В контроле экспрессии генов могут участвовать амплификация, генные перестройки, переключение классов и посттранскрипционные модификации.

3
Аттенуация транскрипции приводит к преждевременной терминации синтеза некоторых молекул РНК. Феномен аттенюации транскрипции изучали преимущественно на бактериях, где с помощью этого механизма регулируется экспрессия многих генов. Для осуществления регуляции такого типа в начале РНК-цепи должна присутствовать определенная последовательность нуклеотидов, которая позволяла бы молекуле РНК находиться в одной из двух возможных конформаций. Более стабильную конформацию имеет петля РНК, которая служит сигналом терминации для бактериальной РНК-полимеразы; в результате синтез РНК преждевременно останавливается. Однако, если с определенной последовательностью растущей молекулы РНК связывается регуляторная молекула, цепь РНК приобретает такую конформацию, при которой сигнала терминации не образуется, а получается длинная функциональная молекула РНК.У бактерий таким регуляторным компонентом обычно служит рибосома, которая в процессе трансляции «садится» на растущую цепь РНК. У эукариот аттенюация транскрипции участвует в регуляции небольшого числа генов. Так как в клеточном ядре функционально активные рибосомы отсутствуют, возможно, что с определенными последовательностями РНК связываются регуляторные молекулы, однако механизм аттенюации в клетках эукариот не изучен.

4
Регуляция экспрессии генов с помощью аттенюации транскрипции у прокариот. По мере роста РНК- транскрипта за счет добавления нуклеотидов к 3'-концу, РНК-транскрипт приобретает одну из двух указанных информации. Наиболее стабильной является конформация А, которая содержит две шпильки с двойной спиралью, возникшие при спаривании комплементарных оснований. В связи с тем, что за спиралью шпильки, образовавшейся при спаривании сайтов 3 и 4, следует серия U- нуклеотидов, шпилька служит сигналом терминации для бактериальной РНК-полимеразы. Конформация Б образуется, если с сайтом 1 на транскрипте РНК связывается регуляторный белок или рибосома, в результате свободный сайт 2 спаривается с сайтом 3, при этом сигнал терминации пропадает и образуется длинный функциональный транскрипт РНК, который и запускает экспрессию гена.

5
Посттранскрипционный контроль Посттранскрипционный контроль генов позволяет системе регуляции суточных ритмов задавать 24-х часовой ритм и обеспечивать устойчивость к резким изменениям, которые могли бы разрушить систему. Критическая форма регуляции всех генов – инициация транскрипции. Посттранскрипционный контроль осуществляется после того, как РНК- полимераза связалась с промотором гена и начала синтез РНК.

6
Есть 4 посттранскрипционных механизма в работе биологических часов. Альтернативный сплайсинг РНК. Регуляция происходит на уровне мРНК, а не на уровне транскрипции. Основные моменты этого уровня регуляции были продемонстрированы на Drosophila. У Drosophila обнаружено два варианта мРНК гена per, которые возникают в результате альтернативного сплайсинга по 3' нетранслируемой области. Т.к. альтернативный сплайсинг касается нетранслируемой области транскриптов, белки, считываемые с двух вариантов мРНК идентичны. Контроль трансляции. При транскрипции и дальнейшей трансляции проявляются задержка между пиковыми уровнями мРНК гена и соответствующего белка, чтобы позволить мРНК накапливаться прежде, чем ген подавляется отрицательной обратной связью. Точные механизмы, которые вызывают задержки неясны.

7
Транслокация в ядро. Контроль происходит на уровне переноса из цитоплазмы в ядро белка - отрицательного компонента петли. Такие белки в дополнение к PAS - домену содержат сигналы ядерной и цитоплазматической локализации. Домены цитоплазматической локализации удерживают белки в цитоплазме. При гетеро димеризации эти домены оказываются скрытыми и гетеро димер может быть перенесен в ядро. Во многих случаях механизм, с помощью которого отрицательные элементы переносятся в ядро, неясен. Деградация белка. Все белки в любых биологических часах имеют короткий период полужизни, что препятствует их накоплению, пока не будет достигнуто равновесие. Таким образом видно, что контроль генов, регулирующих суточный ритм осуществляется не только на уровне транскрипции, но и на посттранскрипционном уровне.

8
Процессинг РНК (посттранскрипционные модификации РНК) Процессинг РНК (посттранскрипционные модификации РНК) совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК. В зависимости от типа РНК (матричные, рибосомные, транспортные, малые ядерные) их предшественники подвергаются разным последовательным модификациям. Например, предшественники матричных РНК подвергаются кэппированию, сплайсингу, полиаденилированию, метилированию и иногда редактированию.
 Процессинг РНК (посттранскрипционные модификации РНК) совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК. В зависимости от типа РНК (матр")
9
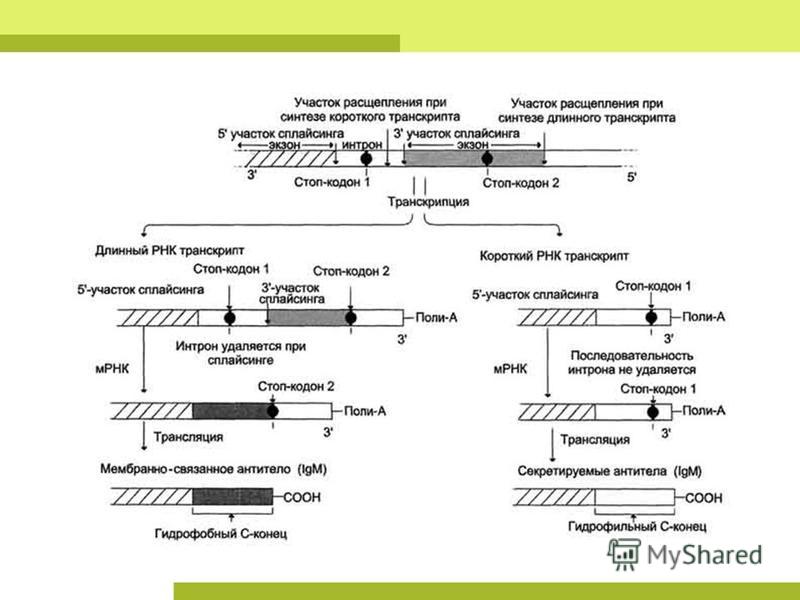
Процессинг РНК стадия отличающая реализацию генетической информации у про- и эукариот. ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция) пространственно не отделён от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полимеразой. Прокариотические мРНК часто полицистронные, то есть содержат несколько независимых генов. ЭУКАРИОТЫ. мРНК эукариот синтезируется в виде предшественника, пре- мРНК, претерпевающего затем сложное стадийное созревание процессинг, включающий присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к её 3'- концу (полиаденилирование), выщепление незначащих участков интронов и соединение друг с другом значащих участков экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
 пространственно не отделён от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полим")
10
Кэпирование Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина через необычный для РНК 5',5'- три фосфатный мостик, а также метилирование остатков рибозы двух первых нуклеотидов. Процесс копирования происходит во время синтеза молекулы пре-мРНК. Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 53'. Функции капа и связанных с ним белков: участие в сплайсинге; участие в процессинге 3'-конца мРНК; экспорт мРНК из ядра; защита 5'-конца транскрипта от экзонуклеаз; участие в инициации трансляции.

11
Этапы процессинга

12
Полиаденилирование Фермент поли(А)-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'- AAUAAA-3' на 3'-конце транскрипта, за которой следует 5'-CA-3'. Вторая последовательность является сайтом разрезания. Сплайсинг После полиаденилирования мРНК подвергается сплайсингу, в ходе которого удаляются интроны (участки, которые не кодируют белки), а экзоны (участки, кодирующие белки) сшиваются и образуют единую молекулу. Сплайсинг катализируется крупным нуклеопротеидным комплексом сплайсосомой, состоящей из белков и малых ядерных РНК. Многие пре-мРНК могут быть подвергнуты сплайсингу разными путями, при этом образуются разные зрелые мРНК, кодирующие разные последовательности аминокислот (альтернативный сплайсинг).
-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'- AAUAAA-3' на 3'-конце транскрипта, за которой с")
13
Редактирование Редактирование РНК процесс, в ходе которого информация, содержащаяся в молекуле РНК, изменяется путём химической модификации оснований. Метилирование мРНК эукариот подвергаются посттранскрипционному метилированию. Наиболее распространённой модификацией является метилирование остатков аденозина по положению N6 с образованием N6- метиладенозина (m6A). Этот процесс катализируют ферменты N6- аденозинметилтрансферазы, которые распознают остатки аденозина в консенсусных последовательностях GAC (70 % случаев) и AAC (30 % случаев). Соответствующие деметилазы катализируют обратный процесс деметилирования. Учитывая обратимость и динамичность процесса метилирования мРНК, а также повышенную концентрацию m6A в длинных экзонах и вокруг стоп-кодонов, предполагают, что метилирование мРНК выполняет регуляторную функцию.

14
Изменение стабильности мРНК Для того, чтобы участвовать в синтезе белка, мРНК должна выйти из ядра в цитоплазму через ядерные поры. Установлено, что в ядре клеток обычно синтезируется больший набор гетерогенных РНК, чем тот, что выходит в цитоплазму. Многие продукты транскрипции подвергаются расщеплению нуклеазами, а те мРНК, что, транспортируются из ядра в цитоплазму, защищаются от гидролитического разрушения, образуя комплексы с белками. Время жизни эукариотических мРНК значительно больше (t 1/2 составляет от нескольких часов до нескольких дней), чем t 1/2 мРНК прокариотов, равное нескольким минутам. Очевидно, что стабильность молекул мРНК - фактор, изменение которого влияет на уровень трансляции. Стабилизация мРНК при фиксированной скорости транскрипции приводит к накоплению и увеличению количества образующегося белкового продукта. Продолжительность жизни разных мРНК варьирует в достаточно широких пределах. Некоторые гены кодируют продукт с большой продолжительностью жизни. Так, в ходе транскрипции гена β-глобина образуется мРНК с t 1/2, равной примерно 10 ч. Другие гены образуют мРНК с короткой продолжительностью жизни: мРНК, на которых синтезируются факторы роста,

16
Рнк-интерференция Пострантранскрипционная регуляция – это процессы, которые происходят после завершения траскрипции – синтеза матричной РНК (мРНК) на ДНК – матрице. В дальнейшем мРНК может быть использована клеткой либо для синтеза белков, либо для деградации (например, её избыточного количества). Один из механизмов деградации мРНК осуществляется с участием микроРНК. В настоящее время известно несколько сотен микроРНК, объединенных в целые семейства в зависимости от свое функциональной активности как в норме так и специфично при ряде заболеваний. МикроРНК являются некодирующими разновидностями РНК и способны связываться с целевыми мРНК – мишенями (одной или несколькими). Механизм РНК-РНК связывания на комплиментарной основе с последующей деградацией с участием набора белковых комплексов носит также название РНК- интерференции. РНК-интерференция – это торможение экспрессии гена на стадии трансляции или транскрипции действием малых РНК. Они выключают работу тех генов, последовательности которых отвечают цепи молекулы малой РНК.
 на ДНК – матрице. В дальнейшем мРНК может быть использована клеткой либо для синтеза белков, либо для д")
17
Деградация мРНК. Время жизни эукариотических мРНК составляет от нескольких часов до нескольких дней, а прокариотических – несколько минут. Продолжительность жизни мРНК является важным механизмом регуляции синтеза белков. Разрушение мРНК у эукариот осуществляется 3-РНК-азами (у прокариот 5- РНК-азами). мРНК эукариот начинает разрушаться с некодирующего полиаденилатного фрагмента. Этот фрагмент подобно теломерным последовательностям ДНК служит буферной зоной, которая защищает кодирующую часть мРНК от разрушения. В промежутке между завершением трансляции мРНК одной рибосомой и началом трансляции следующей 3- РНК-аза успевает отщепить от ее поли(А) фрагмент нуклеотидов. Когда в этом фрагменте остается около 50 нуклеотидов (критическая длина), быстро наступает полный гидролиз мРНК. Кратность трансляции мРНК обычно не превышает раз. Разница в продолжительности жизни различных мРНК объясняется как разной длиной поли-(А) -фрагмента, так и тем, что в короткоживущих мРНК между кодирующей частью и полиаденилатным фрагментом находятся АУ-багатые элементы. Наличие последовательностей обогащенных аденином и урацилом резко усиливает деградацию поли(А) - фрагмента.

18
Спасибо за внимание!

Еще похожие презентации в нашем архиве:




 Оперон (Ф.Жакоб, Ж.Л. Моно, 1961 г.) – группа генов, кодирующих белки,")

 - биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом.")













