Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Әл – Фараби атындағы Қазақ ұлттық университеті Биология және Биотехнология факультеті Репликация процесінің жалпы бейнесі. Мезельсон және Сталь тәжірибесі. Репликацияның жартылай консервативті механизмі. Орындаған: ББ топ Тексерген: Чунетова Ж.Ж.

2
Репликация деп - ДНҚ -нің екі еселенуін айтады. Репликацияның нәтижесінде бір ДНҚ ден сондай екі ДНҚ пайда болады. Уотсон мен Крик тің гипотезасы бойынша ДНҚ нің қос спиралінің әрбір тізбегі репликация кезінде пайда болатын комплементарлық жаңа тізбектер үшін матрица болып табылады. Жаңадан пайда болатын екі ДНҚ молекуласының әрқайсысы ескі ДНҚ молекуласы секілді қос спиральді. Оның себебі ескі ДНҚ қос спиралінің екі тізбегі ажырағаннан кейін,әрбір тізбектердің бойына соларға арқылы ескі тізбекпен сутегілік байланыспен комплементарлық спираль түзейді. Сонымен жаңа қос спиральді ДНҚ нің бір тізбегі жаңадан болса, екінші тізбегі ескі болады. ДНК репликациясы

3
ДНК репликациясының моделі

4
ДНК репликациясы клетканың бөлінуі кезіндегі ең маңызды кілт. Негізінде, ДНК- ның екі еселенуі толық бөлініп болғанда, тек бір рет жүреді. ДНК екі еселенуінің реттелуі механизмдерімен айқын анықталады. Репликация 3 деңгейде өтеді: Репликация инициациясы Репликация инициациясы Элонгация Элонгация Репликация терминациясы Репликация терминациясы

5
1. Репликация инициациясы – бұл ДНК молекуласы репликациясының (екі еселенуі) ең бастапқы фазасы, яғни матрицадағы ескі екі еселенген ата- аналық молекуласының ДНК жіпшелеріндегі екі жаңа туындысының пайда болуы. Репликация деңгейі
 ең бастапқы фазасы, яғни матрицадағы ескі екі еселенген ата- аналық молекуласының ДНК жіпшелеріндегі екі жаңа туындысының пайда болуы. Репликация деңгейі")
8
2. Элонгация – аминқышқылдарын артынша қосу бойынша полипептидті тізбекті өсіре отырып, 5- тан 3 бағытындағы иницирлеуші кодонмен жүретін, нуклеотидтердің триплеттер түрінде аминоацил-тРН көмегімен мРНК – ны оқитын рибосома. Кодонды нақты тану үшін А аймағында аминоацил-тРНК сәйкес қосылуын талап етеді. Аминоацил- тРНК молекуласы және ГТФ жүйесін EF1 элонгация факторы қалыптастырады. Осының әсерінен аминоацил-тРНК рибосомаға қосылуы мүмкін. Фосфат және EF1-ГДФ жүйесінің босап шығуы жүреді. EF1-ГДФ жүйесі ГТФ және басқада бос белоктық факторлардың қатысуымен қайта EF1-ГТФ-қа айналады.

10
3. Терминация – мРНК кодоны атауланған табыс кезінде полиппептидті тізбектің тоқтауы; сондай-ақ терминация –ДНК-ның репликация процесі кезінде немесе РНК синтезінің транскрипция процесінің аяқталуы.

12
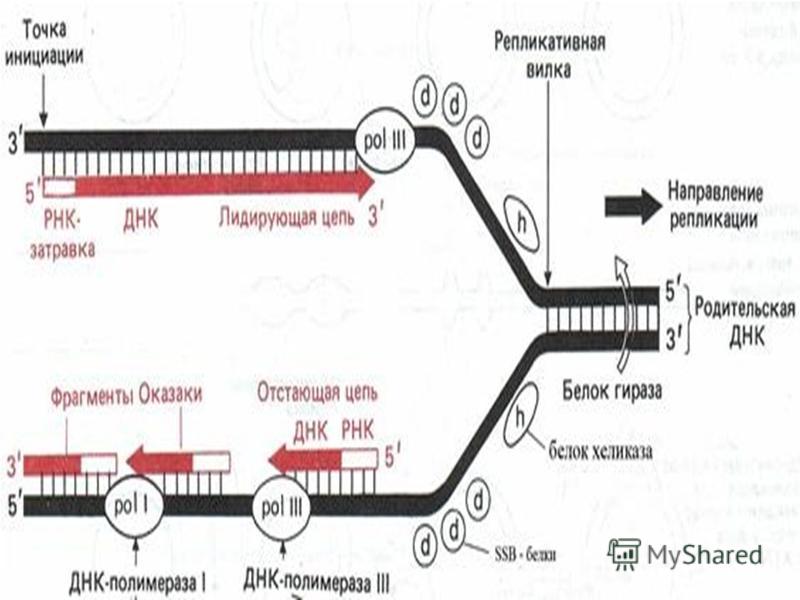
Клетканың репликациясы ДНК – ның спецификалық нүктесінен басталады (репликацияның басталу аймағы) және жүруі екі бағытта жалғасады. Соның нәтижесінде екі репликативті вилка пайда болады. Олар қарама-қарсы бағытта қозғалады, яғни екі тізбекте бір мезгілде репликацияланады.
 және жүруі екі бағытта жалғасады. Соның нәтижесінде екі репликативті вилка пайда болады. Олар қарама-қарсы бағытта қозғалады, яғни екі тізбекте бір мезг")
13
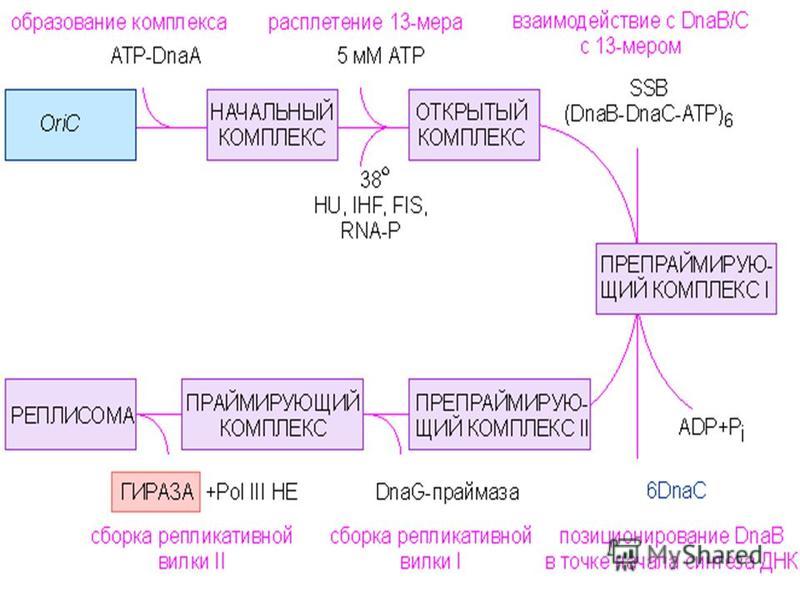
1) Аналық ДНҚ-ны репликация процесіне дайындайтын белоктар 2) Полимеризация процесін іске асыратын ферменттер 3) ДНҚ-ң репликациясын аяқтайтын ферменттер Репликация процесіне қатысатын белоктарды 3 топқа бөлінеді:
 Аналық ДНҚ-ны репликация процесіне дайындайтын белоктар 2) Полимеризация процесін іске асыратын ферменттер 3) ДНҚ-ң репликациясын аяқтайтын ферменттер Репликация процесіне қатысатын белоктарды 3 топқа бөлінеді:")
14
Репликация процесі ДНҚ полинуклеотидтігінің бірнеше нүктелерінде басталады. Осы нүктелерге оларды танитын ерекше белоктар байланысады. Бактериялар бұл белоктары DnaA белоктар деп аталады, репликацияны ынталандыратын бірінші белоктар. Репликация іске асатын ДНҚ-ның аяғында қосарланған спираль ажырап бір тізбекті бөліктері түзілу керек. Спиральдің ажырауы геликаза ферментінің қатысуымен іске асады. Спираль бөліну үшін АТФ-тың энергиясы жұмсалады. Аналық ДНҚ-ны репликация процесіне дайындайтын белоктар

15
Эукариоттарда 5 ДНҚ-полимераза белгілі:α және δ-полимеразалар ядролық ДНҚ-ның репликациясына,γ-полимераза митохондрия репликациясына, β және ε-полимеразалар ДНҚ-ның репарациясына қатысады

16
Полинуклеотидтік тізбектің түзілуіне қатысатын негізгі фермент ДНҚ-полимераза. Эукариоттарда 5 ДНҚ-полимераза белгілі:α және δ-полимеразалар ядролық ДНҚ-ның репликациясына,γ-полимераза митохондрия репликациясына, β және ε-полимеразалар ДНҚ-ның репарациясына қатысады. ДНҚ-полимеразаларды ДНҚ-ның молекуласымен PCNA-белок байланыстырады. Полинуклеотидтік тізбектің синтезі басталу үшін, алдымен қысқа полинуклеотидтік тізбек түзіледі, оны РНҚ-заставка немесе праймер деп аталады. РНҚ-праймаза праймердің синтезін катализдейді. РНҚ-праймазаны ерекше белок активтендіреді-праймазаның активаторы. Полимеризация процесін іске асыратын ферменттер

17
ДНҚ-ның полинуклеотидтік тізбектері өзара тығыз орналасқан фрагменттер түрінде түзіледі. Осы нуклеотидтік фрагменттерді ДНҚ-лигаза өзара тігеді. Репликация барысында түзілген жаңа полинуклеотидтік тізбектердің барысында праймер орналасады. Полинуклеотидтік тізбегінің синнтезі аяқталғаннан кейін праймер бөлініп кетеді, сондықтан жаңа тізбектер праймердің ұзындығына қысқарған болады. Бұл аналық ДНҚ молекуласының бөліктерін теломерлы аймақтар деп аталады. ДНҚ молекуласының репликациясы толық аяқталу үшін осы теломерлы бөліктерінің репликациясы іске асуы қажет. Праймерлер бөлініп кеткеннен кейін теломераза ферментінің қатысуымен ДНҚ-ның полинуклеотидтік тізбектерінің ұзындығы аналық ДНҚ-ның полинуклеотидтік тізбегінің ұзындығына дейін ұзарады. ДНҚ-ң репликациясын аяқтайтын ферменттер

18
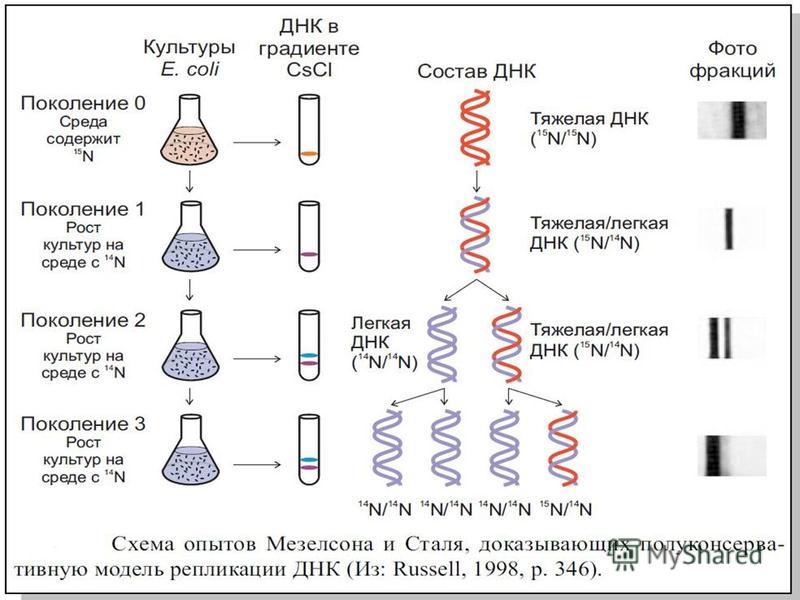
Мезельсон және Сталь тәжірибесі Мезельсон және Сталь тәжірибесі – 1958 жылы Сталь Франклин мен Мэтью Мезельсон атты екі молекулалық биологтармен жүргізілген тәжірибе. Олар, ДНК – ның екі еселенуін жартылай консервативті сипат алады. Бұл дегеніміз әр екі еселенген спираль ДНК бір ескі (матрицалық)тізбектен және бір синтезделген тізбектен тұрады.

19
Сталь Франклин Мэтью Мезельсон

20
М. Мезельсон мен Ф.Сталь 1958 жылы жартылай консервативті моделдің барын дәлелдеді. Олар E.coli бактериясын бірнеше буындар да N 15 атомы азотты хлорид аммонидің қорашаған ортаның жалғыз көзі. Нәтижесінде барлық клеткалық құрылымдар ауыр азот N 15 бактерияның құрамында болған.

22
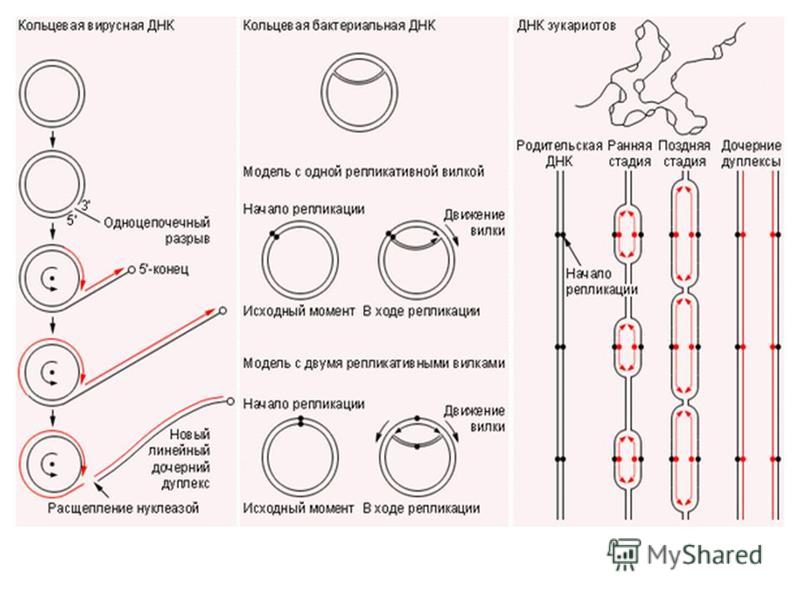
ДНҚ репликациясы молекуланың кез-келген кездейсоқ нүктесінен емес, репликацияның басталу нүктелері деп аталатын арнайы орындардан басталады. ДНҚ репликациясының механизмі туралы қазіргі көзқарастарды қарастырғанда репликонда жүретін бұл процестің бір қатар кезеңдерін бөліп қарауға болады. Бірінші кезең ДНҚ молекуласының екі полинуклеотиды жіпшелерінің жұмыс істеп тұрған репликоның шекарасында жылдам спиралдануына және олардың комплеметарлы негіздер жұптарының арасындағы сутекті байланыстардың бұзылуы арұылы бөлініп кетуіне байланысты болады. Екінші кезеңде жаңа нуклеоидты тізбектің комплеметарлық матрицалық рекция жүреді. Бұл процес бірнеше тұрлы ДНҚ- полимераза ферментерінің көмегімен жаңа тізбектерді нуклеоидтердің полимеризациясы дақылы жүзеге асады Репликация механизмдері

24
ДНК репликация – ДНК синтезі – жартылай консервативті механизм юойынша жүреді. Уотсон мен Крикті гипотезесына сәйкес, әр тізбектің екі тал комплементарлы екі еселенген репликация комплементарлы аталған тізбектер. Осыдан екі еселенген ДНК-ның екі тізбекті молекуласы, ата- анасының ДНК – мен бірдей. ДНК – ның қайта синтезделген тізбегінің біреуі, ата – ананың бір өзгермейтін тізбегінде осы молекулалар кездеседі. Репликацияны ң жартылай консервативті механизмі

26
1.Репликация жартылай консервативтi әдiспен жүредi. Ескi ДНҚ-ның әр тiзбегiн қалып ретiнде пайдаланып, ДНҚ–полимераза ферментi комплементарлық принцип бойынша бос нуклеотидтерден екiншi тiзбектi түзедi. Ол үшін тiзбектер арасындағы сутектiк байланыстар үзіліп, ДНҚ-ның екi тiзбегі бiр бiрiнен ажырап, босаңсып,репликация айырын түзедi. Эукариоттардағы ДНҚ репликациясының ерекшелiктерi:

28
2. ДНҚ молекуласының аса ұзын болуына байланысты репликация бiрден бiрнеше жерден басталып (полирепликонды түрде), екi бағытта келесi репликативтiк айырмен кездескенше жүредi. Репликацияның басталатын нүктелері нуклеотидердiң арнайы ретiмен анықталыпинициация нүктесi деп аталады. Олардың саны әр хромосоманың ДНҚ-да нақты белгілі болады. Репликацияның басталу нүктесiнен келесi репликация айырымен кездесетiн жерге дейiнгi ДНҚ-ның бөлiгi репликон деп аталады - бұл репликация бiрлiгi. Прокариоттар мен органоидтардағы (митохондриялар мен пластидтер) ДНҚ молекуласының хромосомалық ДНҚ-нан айырмашылығы - оларда тек бiр инициация нүктесi болады, сондықтан олар бiр репликон деп саналады.
, екi бағытта келесi репликативтiк айырмен кездескенше жүредi. Репликацияның басталатын нүктелері нуклеотидердiң арнайы ретiмен анықталып")
29
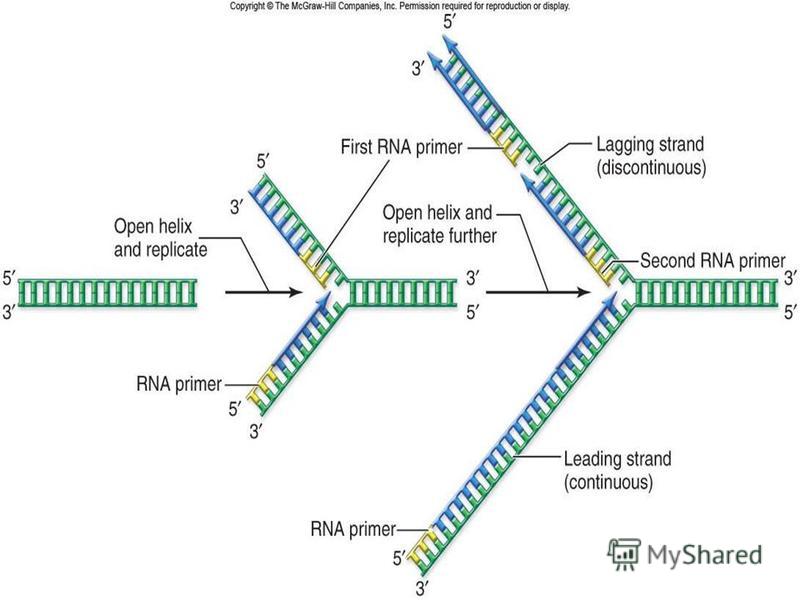
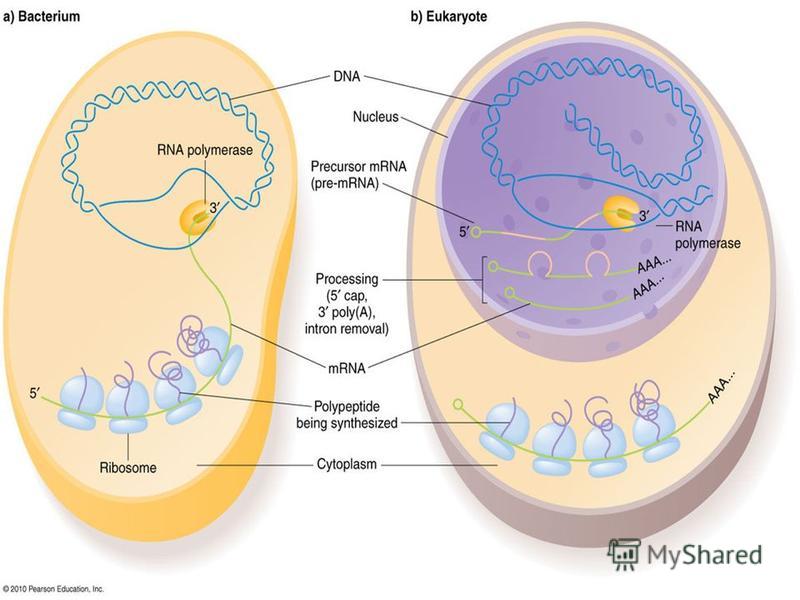
3. Жасушадағы ДНҚ-ның репликациясы жасушалық циклдың S-кезеңiнде жүредi. Бiрақ, репликондардың репликациясы бiркелкi емес, асинхронды түрде жүредi. Мысалы, рРНҚ туралы ақпараты бар ДНҚ бөлiктерi S- кезеңiнiң басында екi еселенедi де, басқа бөлiктерi кейiн еселенедi. Митохондриялық ДНҚ-ның репликациясы көбiнесе жасушаның әрбір бөлiнуі алдында, G 2 -кезеңiнде, болып кетедi. Ал бөлiнбейтiн жасушаларда (мысалы бауыр жасушалары) митохондириялық ДНҚ-ның репликациясы физиологиялық ескіруге байланысты митоздық циклдың фазаларына тәуелсiз жүреді. 4. НҚ синтезі тек 5 -3 бағытта жүретiн, ал ДНҚ тiзбектерiнiң қарама қарсы (антипаралельді) болғандықтан, ДНҚ-ның бiр тiзбегi үздiксiз түзiлiп, лидерлiк деп, екiншiсi соңынан бiр тiзбекке жалғанатын кішірек үзінділер (Оказаки фрагменттерi) түрiнде синтезделіп, iлесушi тізбек деп аталады. Жалпы ДНК репликациясы про - және эукариоттарда ұксас, 6ipaK эукариоттарда синтез жылдамдыгы бірқатар төмен (1 сек нуклеотид шамасында) болады, прокариоттарда (1 сек нуклеотид шамасында) жылдамырак жүреді. Ce6e6i, эукариоттың ДНҚ-сы акуызбен бepiK байланысқан, ол оның деспиральдануын тежейді, репликациясын баяулатады.

31
Назарларыңызға рахмет!!!

Еще похожие презентации в нашем архиве:















")

 енгізді, оның теориясын А.В.Гадолин.")

 аксонометрия осьтерін қалай жүргізеді? 2) аксонометрия.")



