Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Биосинтез белка Основные вопросы: 1. Транскрипция 2. Трансляция 3. Общая схема б-за белка

3
РНК получает наследственную информацию Прежде чем начнут синтезироваться белки, информацию об их строении необходимо "достать" из ДНК и доставить ее к месту синтеза белков. Этим занимаются информационные или матричные РНК. Одновременно клетке нужны транспортеры аминокислот – транспортные РНК и структурные компоненты органелл, синтезирующих белок, – рибосомальные РНК. Вся информация о строении транспортных и рибосомальных РНК также находится в ДНК. Поэтому существует процесс переписывания или транскрипции данных с ДНК на РНК (англ. transcription – переписывание) – биосинтез РНК на матрице ДНК.

5
Транскрипция - первая стадия реализации генетической информации в клетке. В ходе процесса образуются молекулы мРНК, служащие матрицей для синтеза белков, а также транспортные, рибосомальные и другие виды молекул РНК, выполняющие структурные, адаптерные и каталитические функции (рис. 4-26).

6
Транскрипция у эукариотов происходит в ядре. В основе механизма транскрипции лежит тот же структурный.принцип комплементарного спаривания оснований в молекуле РНК (G C, A=U и Т=А). ДНК служит только матрицей и в ходе транскрипции не изменяется. Рибонукле-озидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ) -субстраты и источники энергии, необходимые для протекания полимеразной реакции, образования 3',5'-фосфодиэфирной связи между рибонуклеозидмонофосфатами. Синтез молекул РНК начинается в определённых последовательностях (сайтах) ДНК, которые называют промоторы, и завершается в терминирующих участках (сайты терминации). Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции - транскриптон. У эукариотов в состав транскриптона, как правило, входит один ген (рис. 4-27), у прокариотов несколько. В каждом транскриптоне присутствует неинформативная зона; она содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
. ДНК служит только матрицей и в ходе транскрипции не изменяется. Рибон")
7
Транскрипционые факторы - белки, взаимодействующие с определёнными регуляторными сайтами и ускоряющие или замедляющие процесс транскрипции. Соотношение информативной и неинформативной частей в транскриптонах эукариотов составляет в среднем 1:9 (у прокариотов 9:1).

8
Соседние транскриптоны могут быть отделены друг от друга нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью индивидуальное считывание (транскрипцию) разных генов.
 разных генов.")
9
В каждом транскриптоне транскрибируется только одна из двух цепей ДНК, которая называется матричной, вторая, комплементарная ей цепь, называется кодирующей. Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда антипараллельна синтезируемой нуклеиновой кислоте (рис. 4-28). Транскрипция не связана с фазами клеточного цикла; она может ускоряться и замедляться в зависимости от потребности клетки или организма в определённом белке.

10
РНК-полимеразы Биосинтез РНК осуществляется ДНК-зависимыми РНК- полимеразами. В ядрах эукариотов обнаружены 3 специализированные РНК- полимеразы: РНК-полимераза I, синтезирующая пре-рРНК; РНК-полимераза II, ответственная за синтез пре-мРНК; РНК-полимераза III, синтезирующая пре-тРНК. РНК-полимеразы - олигомерные ферменты, состоящие из нескольких субъединиц - 2α, β, β', σ. Субъединица о (сигма) выполняет регуляторную функцию, это один из факторов инициации транскрипции, РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.

11
Стадии транскрипции Инициация Активация промотора происходит с помощью большого белка - ТАТА-фактора, называемого так потому, что он взаимодействует со специфической последовательностью нуклеотидов промотора - ТАТААА- (ТАТА-бокс) (рис. 4-29).
 (рис. 4-29).")
12
Присоединение ТАТА-фактора облегчает взаимодействие промотора с РНК-полимеразой. Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК (рис. 4-30). После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков, σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.

13
1 - присоединение ТАТА- фактора к промотору. Чтобы промотор был узнан РНК-полимеразой, необходимо образование транскрипционного комплекса ТАТА-фактор/ТАТА-бокс (промотор). ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегчает использование промотора многими молекулами РНК- полимеразы; 2 - образование транскрипционной вилки; 3 - элонгация; 4.- терминация.
. ТАТА-фактор остаётся связанным с ТАТА-боксом во время транскрипции, это облегч")
15
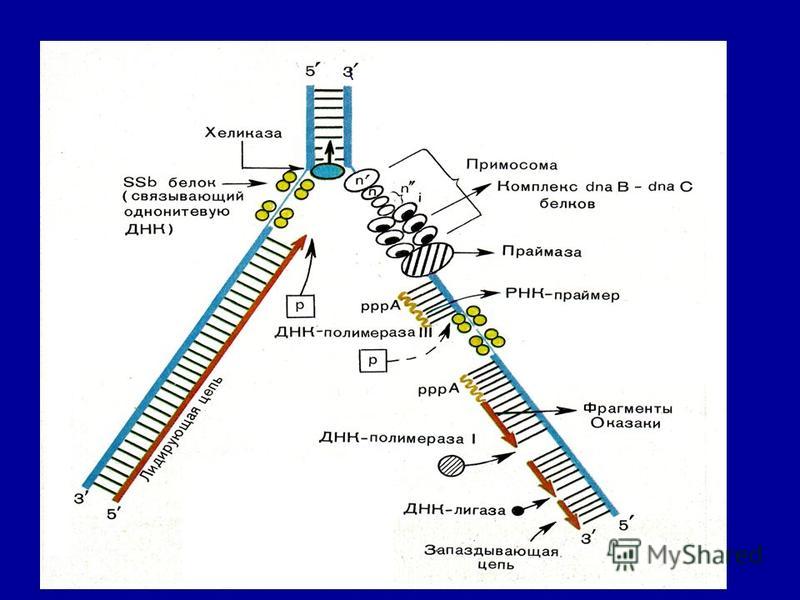
Элонгация Факторы элонгации повышают активность РНК- полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК.

16
Терминация Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы - терминаторах (сайты терминации). Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
. Фактор терминации облегчает отделени")
17
Ф гтф GF Т А Ц Г ГЦ. АТ. ф АУГЦ Для начала транскрипции необходимы: фермент РНК-полимераза, фактор транскрипции (GF) и ГТФ. Фермент связывается с GF и присоединяется к промотору – специальному участку ДНК. К этому комплексу присоединяется ГТФ и происходит раскручивание ДНК. Фермент двигается вдоль одной из цепей ДНК, присоединяя комплементарные нуклеотиды, образующейся иРНК. Дойдя до терминатора, РНК-полимераза присоединят TF и отщепляется от ДНК. Транскрипция TF и РНК
 и ГТФ. Фермент связывается с GF и присоединяется к промотору – специальному участку ДНК. К этому комплексу присоединяется ГТФ и проис")
18
Ковалентная модификация (процессинг) матричной РНК Первичные транскрипты мРНК, прежде чем будут использованы в ходе синтеза белка, подвергаются ряду ковалентных модификаций. Эти модификации необходимы для функционирования мРНК в качестве матрицы.
 матричной РНК Первичные транскрипты мРНК, прежде чем будут использованы в ходе синтеза белка, подвергаются ряду ковалентных модификаций. Эти модификации необходимы для функционирования мРНК в качестве матрицы.")
19
Кэпирование (англ. cap – шапка) Когда длина первичного транскрипта достигает примерно 30 нуклеотидных остатков, происходит копирование Процесс состоит в присоединении к 5'-трифосфату концевого нуклеотида пре-мРНК 5'-углерода N7-метил-гуанозина. "Кэп" необходим для защиты молекулы РНК от действия 5'- экзонуклеаз в цитоплазме., а также для связывания мРНК с рибосомой и для начала трансляции. Кэпирование необходимо для инициации синтеза белка, так как инициирующие триплеты AUG, GUG распознаются рибосомой только если присутствует кип.
 Когда длина первичного транскрипта достигает примерно 30 нуклеотидных остатков, происходит копирование Процесс состоит в присоединении к 5'-трифосфату концевого нуклеотида пре-мРНК 5'-углерода N7-метил-гуанозина.")
20
Полиаденилирование Полиаденилирование – при помощи полиаденилат- полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100 до 200 адениловых нуклеотидов, формирующих полиадениловый фрагмент (поли(А)-хвост). Поли(А)-хвост необходим для защиты молекулы РНК от экзонуклеаз, работающих с 3'-конца.
-хвост). Поли(А)-хвост")
21
Транскрипция Для экспрессии гена, т.е. синтеза закодированных в нем белков, последовательность нуклеотидов кодирующей цепи ДНК должна быть трансформирована в аминокислотную последовательность. Как в любом матричном биосинтезе в транскрипции выделяют 5 необходимых элементов: 1. матрица – одна из цепей ДНК, 2. растущая цепь – РНК, 3. субстрат для синтеза – рибонуклеотиды (УТФ, ГТФ, ЦТФ, АТФ), 4. источник энергии – УТФ, ГТФ, ЦТФ, АТФ. 5. ферменты РНК-полимеразы и белковые факторы транскрипции

22
Транскрипция Биосинтез РНК происходит в участке ДНК, который называется транскриптон, с одного края он ограничен промотором (начало), с другого – терминатором (конец). РНК-полимеразы эукариот имеют по две больших субъединицы и несколько малых субъединиц Поскольку ДНК не принимает непосредственного участия в синтезе белка, информация, хранящаяся в ядре, должна быть перенесена на рибосомы, где собственно и осуществляется биосинтез белков. Для этого соответствующий участок кодирующей цепи ДНК считывается (транскрибируется) с образованием гетерогенной ядерной РНК [гяРНК (hnRNA)], т. е. последовательность этой РНК комплементарна кодирующей цепи ДНК. Поскольку в РНК вместо тимина содержится урацил, AAG триплет ДНК трансформируется в UUC-кодон гяРНК
, с другого – терминатором (конец). РНК-полимеразы эукариот имеют по две больших субъединицы и несколько малых субъеди")
23
Ген эукариот содержит наряду с кодирующими (экзоны) также некодирующие (интроны) последовательности. Фермент РНК-полимераза катализирует транскрипцию как экзонов, так и интронов с образованием гетерогенной ядерной РНК (гяРНК), называемой также первичным транскриптом. Термин «интроны» означает вставочные, не транслирующие последовательности нуклеотидов в ДНК эукариот.
 также некодирующие (интроны) последовательности. Фермент РНК-полимераза катализирует транскрипцию как экзонов, так и интронов с образованием гетерогенной ядерной РНК (гяРНК), называемой также первич")
24
Процессинг. Созревание РНК-Сплайсинг Между транскрипцией и трансляцией существует еще одного важное звено – образование «зрелой» молекулы мРНК. Этот этап получил название процессинга, или созревания, мРНК, (посттранскрипционные модификации РНК) Процессинг мРНК включает три основных процесса: 1) сплайсинг – удаление некодирующих иностранных последовательностей из мРНК и сшивание образующихся экзонов;
 Проце")
25
Созревание РНК. У эукариот гяРНК, прежде, чем покинуть ядро в виде матричной РНК (мРНК), претерпевает существенные изменения: из молекулы вырезаются избыточные (некодирующие) участки (интроны), а оба конца транскриптов(экзоны) модифицируются путем присоединения дополнительных нуклеотидов.
, претерпевает существенные изменения: из молекулы вырезаются избыточные (некодирующие) участки (интроны), а оба конца транскриптов(экзоны) модифицируются путем пр")
26
Схематичное представление матричной РНК после процессинга 2).Кэпирование (англ. cap – шапка) – происходит еще во время транскрипции. Процесс состоит в присоединении к 5'- трифосфату концевого нуклеотида пре- мРНК 5'-углерода N7-метил-гуанозина. "Кэп" необходим для защиты молекулы РНК от экзонуклеаз, а также для связывания мРНК с рибосомой и для начала трансляции. 3). Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100 до 200 адениловых нуклеотидов, формирующих полиадениловый фрагмент (поли(А)-хвост). Поли(А)-хвост необходим для защиты молекулы РНК от экзонуклеаз, работающих с 3'-конца.
.Кэпирование (англ. cap – шапка) – происходит еще во время транскрипции. Процесс состоит в присоединении к 5'- трифосфату концевого нуклеотида пре- мРНК 5'-углерода N7-метил-гуанозина.")
27
Трансляция Трансляция. Зрелая мРНК попадает в цитоплазму и связывается с рибосомами, преобразующими полученную информацию в аминокислотную последовательность. Рибосомные РНК выполняют функцию структурного элемента рибосом, а также принимают участие в связывании мРНК и образовании пептидных связей. Механизм преобразования генетической информации основан на взаимодействии кодонов мРНК с транспортной РНК, которая переносит на рибосому аминокислоты, связанные с 3'-концом тРНК, в соответствии с информацией, закодированной в мРНК. Примерно в середине цепи тРНК расположен триплет (например, GAA), антикодон и комплементарный соответствующему кодону а мРНК. Если транслируется кодон UUC, то с ним взаимодействует антикодон в составе Phe-тРНК, несущей на 3'-конце остаток фенилаланина. Таким образом, остаток аминокислоты занимает положение, в котором на него может быть перенесена растущая полипептидная цепь, связанная с соседней тРНК.

28
Генетический код Наследственная информация записана в молекулах НК в виде последовательности нуклеотидов. Определенные участки молекулы ДНК и РНК (у вирусов и фагов) содержат информацию о первичной структуре одного белка и называются генами. 1 ген = 1 молекула белка Поэтому наследственную информацию, которую содержат ДНК называют генетической.
 содержат информацию о первичной структуре одного белка и называются генами. 1 ген =")
29
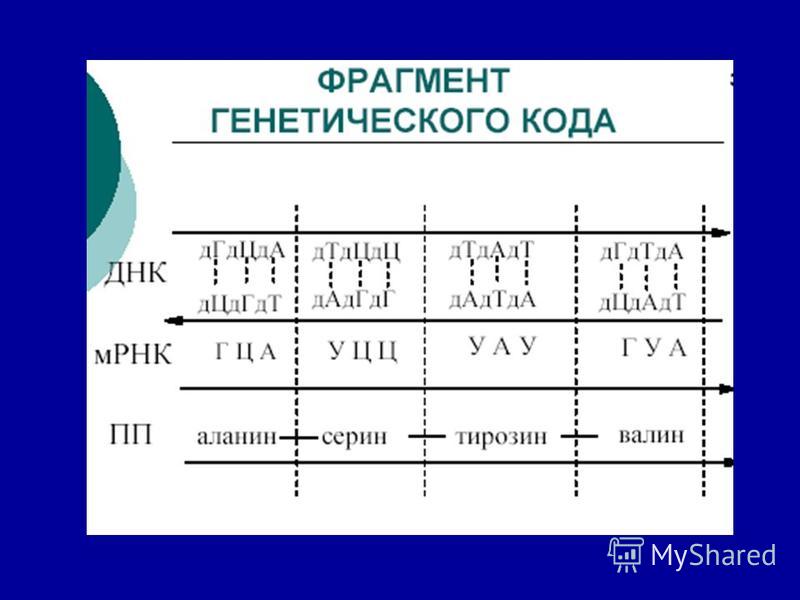
Каждой аминокислоте в полипептидной цепи соответствует комбинация из трех нуклеотидов (триплет) в ДНК Эта зависимость между азотистыми основаниями в ДНК и аминокислотами в белке называется - генетическим кодом.
 в ДНК Эта зависимость между азотистыми основаниями в ДНК и аминокислотами в белке называется - генетическим кодом.")
30
Одна аминокислота закодирована тремя нуклеотидами (один кодон). АЦТ АГЦ ГАТ Триплет, кодон ген АК1 АК2 АК3 белок Пример: АК триптофан закодирована в РНК УГГ, в ДНК - АЦЦ.
. АЦТ АГЦ ГАТ Триплет, кодон ген АК1 АК2 АК3 белок Пример: АК триптофан закодирована в РНК УГГ, в ДНК - АЦЦ.")
31
Имеется 64 кодона: 61 кодон кодирует 20 (21) аминокислот, три кодона являются знаками препинания: кодоны-терминаторы УАА, УАГ, УГА (в РНК). АТЦГАТЦГ 4 3
 аминокислот, три кодона являются знаками препинания: кодоны-терминаторы УАА, УАГ, УГА (в РНК). АТЦГАТЦГ 4 3")
32
Генетический код: Триплетный: каждая аминокислота кодируется триплетом нуклеотидов ДНК и соответствующим кодоном иРНК. Однозначный: один кодон соответствует одной аминокислоте Непрерывный: кодоны мРНК не отделены друг от друга (отсутствуют «запятые») Вырожденный (избыточный): одна аминокислота может кодироваться разными кодонами Не перекрывающийся: каждый нуклеотид в мРНК принадлежит только одному кодону (исключения обнаружены у вирусов). Универсальный: генетический код одинаков для всех организмов. в митохондриях и у инфузорий генетический код имеет некоторые отличия
")
33
эритроциты - двояковогнутые диски, содержат гемоглобин. –Норма: 6-е место – глу –Патология – вал Гемоглобин - белок 1 молекула = 4 полимера 1 полимер = 574 АК При изменении молекулы белка изменяется свойство гемоглобина, возникает наследственное заболевание: серповидно-клеточная анемия. Пример:

35
Синтез белка протекает в несколько стадий 1) Активация аминокислот специфическими синтетазами и их связывание с тРНК. Для синтеза белка необходимо наличие достаточного количества всех аминокислот. Синтез полипептида начинается с 5'-ОН-конца мРНК. 2) Биосинтез пептидной цепи начинается в присутствии инициирующих факторов F1, F2 и F3 (все факторы являются белками), мРНК, инициаторной тРНК, GTP, Mg2+ и рибосомальных субъединиц с коэффициентами седиментации 30S и 50S..
 Активация аминокислот специфическими синтетазами и их связывание с тРНК. Для синтеза белка необходимо наличие достаточного количества всех аминокислот. Синтез полипептида начинается с 5'-ОН-конца мРНК. 2)")
36
3) Образование пептидной связи и рост белковой цепи. Этот процесс происходит в несколько стадий. а) Ориентация отдельных АА-тРНК вдоль мРНК и их взаимодействие с рибосомой (в участке А); Для связывания необходимы GTP и цитоплазматический белок Т. Для того чтобы произошло связывание, в молекуле соответствующей тРНКАА, должен присутствовать антикодон, т.е. триплет оснований с составом, комплементарным триплетному кодону мРНК. б) Образование пептидной связи (или удлинение существующей пептидной цепи на одну аминокислоту под действием пептидилтрансферазы). в) Перемещение пептида, связанного через последнюю тРНК, в участке А рибосомы на участок Р, и освобождение предыдущей тРНК из ее связи с мРНК в участке Р (транслоказа и GTP). В то же время происходит движение мРНК вдоль рибосомы на один кодон.
 Образование пептидной связи и рост белковой цепи. Этот процесс происходит в несколько стадий. а) Ориентация отдельных АА-тРНК вдоль мРНК и их взаимодействие с рибосомой (в участке А); Для связывания необходимы GTP и цитоплазматический белок Т. Для")
37
4) Отделение пептидной цепи от рибосомы происходит в том месте, где на молекуле мРНК встречается «стоп» (или «бессмысленный») кодон (UAA, UAG или UGA). По мере роста цепи она свертывается с образованием вторичной и третичной структур
 Отделение пептидной цепи от рибосомы происходит в том месте, где на молекуле мРНК встречается «стоп» (или «бессмысленный») кодон (UAA, UAG или UGA). По мере роста цепи она свертывается с образованием вторичной и третичной структур")
38
Активация АК Активация свободных аминокислот осуществляется при помощи специфических ферментов – аминоацил-тРНК- синтетаз – в присутствии АТФ. Этот процесс протекает в две стадии: На I стадии аминокислота вступает в реакцию с АТФ, при этом освобождается пирофосфат и образуется активированная АК, которая на II стадии реагирует с соответствующей 3'-ОН- тРНК, и образуется аминоацил-тРНК и освобождается АМФ. Аминокислота присоединяется к свободному концевому 3'- ОН-гидроксилу (или 2'-ОН) АМФ, который вместе с двумя остатками ЦМФ образует концевой триплет ЦЦА, являющийся одинаковым для всех транспортных РНК. Аминоациладенилат Аминоацил-тРНК Аминоацил-тРНК- синтаза Аминоациладенилат АТФPPi Аминоацил-тРНК + тРНК I II

39
Схема биосинтеза Процесс биосинтеза белка состоит из трех стадий: транскрипции (синтез иРНК), сплайсинга («созревание» иРНК) и трансляции (биосинтез первичного белка). Транскрипция и сплайсинг протекают в ядре, а трансляция – в цитоплазме. В трансляции принимают участие тРНК, доставляющие аминокислоты к месту сборки белковой молекулы.
, сплайсинга («созревание» иРНК) и трансляции (биосинтез первичного белка). Транскрипция и сплайсинг протекают в ядре, а трансляция – в цитоплазме. В трансляц")
40
Трансляция После удаления интронов иРНК становится «зрелой» и переносится в цитоплазму. Там с ней связываются рибосомы, которые одна за другой двигаются вдоль иРНК и при участии молекул тРНК синтезируют первичную цепь белка из аминокислот согласно информации, записанной в кодирующем участке иРНК. По прошествии некоторого времени иРНК разрушается под действием ферментов – рибонуклеаз. Первым подвергается деградации хвост poly (А), содержащий нуклеотидов который присоединяется к иРНК в процессе транскрипции и не удаляется при сплайсинге.

41
Субъединицы рибосом присоединяются к иРНК только в процессе трансляции, образуя рибосому, которая продвигается вдоль иРНК и синтезирует первичный белок. После синтеза белка рибосома распадается на субъединицы. Трансляция, как и транскрипция состоит из трех последовательных процессов: инициации (начало синтеза белка), элонгации и терминации.

42
Функциональные участки рибосом Р – пептидильный участок для пептидил-тРНК А – аминоацильный участок для аминоацил-тРНК Е – участок для выхода тРНК из рибосомы Е

43
Первый этап инициации FI иРНК Мет-тРНК На рибосоме имеется два участка: один из них связан с удлиняющейся цепью полипептида (Р-участок), а второй присоединяет новую аминоацил-тРНК.(А-участок) Малая рибосомная субъединица присоединяется к иРНК, а затем к этому комплексу присоединяется инициаторная тРНК (Мет-тРНК) в комплексе с ферментом FI (фактор инициации) и ГТФ, т.к. любой белок начинает синтезироваться с метионина. Инициаторным триплетом служит единственный кодон метионина АУГ- старт-кодон (в некоторых случаях - ГУГ, кот в середине цепи кодирует валин, а в ее начале- метионин) ГТФ РА
, а второй присоединяет новую аминоацил-тРНК.(А-участок) Малая рибосомная субъединица присоединяется к иРНК, а зате")
44
Второй этап инициации АУГ Р А F1 Мет-тРНК Полученный комплекс продвигается по иРНК, пока антикодон УАЦ Мет- тРНК не спарится со старт-кодоном АУГ иРНК. Далее к комплексу (иРНК – малая субъединица – мет-тРНК) присоединяется большая субъединица и образуется комплекс инициации. FI теряет сродство с мет-тРНК и уходит, а мет-тРНК перемещается в Р участок малой субъединицы, в результате чего участок А становится вакантным. В ходе инициации происходит гидролиз ГТФ до ГДФ.
 присоединяется большая субъединица и образ")
45
Инициация трансляции

46
ЭЛОНГАЦИЯ Процесс элонгации включает образование пептидных связей между соседними аминокислотами, при этом очередность присоединения аминокислот определяется очередностью кодонов в иРНК. АУГ ААА ГУЦ ВАЛ МЕТ ЛИЗ После образования инициатор- ного комплекса кодон в молекуле иРНК, следующий за кодоном АУГ, спаривается с комплементарным ему антикодоном соответствующей тРНК, а между карбоксильной группой метионина и аминогруппой следующей аминокислоты с помощью ферментативной активности, присущей большой субъединице, образуется пептидная связь. Метионин отсоединяется от инициаторной тРНК, которая отделяется от рибосомы, «протягивая» последнюю на 1 кодон по иРНК. После образования инициатор- ного комплекса кодон в молекуле иРНК, следующий за кодоном АУГ, спаривается с комплементарным ему антикодоном соответствующей тРНК, а между карбоксильной группой метионина и аминогруппой следующей аминокислоты с помощью ферментативной активности, присущей большой субъединице, образуется пептидная связь. Метионин отсоединяется от инициаторной тРНК, которая отделяется от рибосомы, «протягивая» последнюю на 1 кодон по иРНК.

47
Терминация (освобождение) Элонгация продолжается до тех пор, пока рибосома не дойдет до кодона УАА, УАГ, УГА В конце каждого цистрона расположен один из триплетов (конец цепи), кодирующий момент окончания синтеза полипептида (стоп- кодон, терминирующий кодон, запятые). Белковые факторы терминации (RF) связываются с рибосомой, узнают один из триплетов «конец цепи» и вызывают гидролиз связей между последней тРНК и полипептидной цепью, рибосомой и иРНК. (рибосомная пептидил- трансфераза) В результате чего полипептид освобождается. Рибосома диссоциирует на субъединицы, которые вновь могут участвовать трансляции.
 Элонгация продолжается до тех пор, пока рибосома не дойдет до кодона УАА, УАГ, УГА В конце каждого цистрона расположен один из триплетов (конец цепи), кодирующий момент окончания синтеза полипептида (стоп- кодон, терминирующ")
48
СИНТЕЗ ПЕПТИДНОЙ СВЯЗИ Первая аминокислота цепи (fMet-тРНК) связывается с большой субъединицей рибосомы в участке П. Следующая аминокислота в виде аминоацил-тРНК связывается в участке А рибосомы за счет взаимодействия антикодона тРНК и кодона мРНК. В результате NH2-группа связывающейся аминокислоты оказывается вблизи от карбоксильной группы предыдущей аминокислоты и с помощью пептидилтрансферазы образуется пептидная связь. Образовавшийся дипептид связан с тРНК второй аминокислоты. Далее, дипептид переносится транслоказой, для функционирования которой необходима энергия GTP, из участка А в участок П, вытесняя оттуда освободившуюся тРНК, которая необходима для следующих переносов. Рибосома сдвигается и против А-участка становится следующий кодон. Так восстанавливается исходная ситуация и весь процесс повторяется. Процесс продолжается до тех пор, пока до А участка рибосомы не дойдет «стоп»-кодон (который не кодирует никакой аминокислоты). На этом синтез заканчивается и синтезируемый пептид отделяется от поверхности рибосомы.
 связывается с большой субъединицей рибосомы в участке П. Следующая аминокислота в виде аминоацил-тРНК связывается в участке А рибосомы за счет взаимодействия антикодона тРНК и кодона мРНК. В")
49
На одной иРНК «работают» несколько рибосом. Такой комплекс называется полисома. После завершения синтеза иРНК распадается на нуклеотиды. Весь цикл процессов, связанных с синтезом одной белковой молекулы, занимает в среднем 1-3 с. Полисома из печени содержит 12 рибосом, которые выглядят темными пятнами. А цепочка иРНК на снимке не видна.

50
Аминокислоты

Еще похожие презентации в нашем архиве:





 молекулярной биологии – матричный синтез. Этапы биосинтеза белка: ДНК репликация ДНК транскрипция.")




 Принцип комплементарности и (или м) – РНК, т – РНК, их функции Белки.")







 Аксенова Светлана Джоновна ГОУ СОШ 156 с углубленным изучением информатики Калининского района Учитель биологии Октябрь 2007 г.")




