Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
IMUNITATEA. SISTEMUL IMUN. CELULELE IMUNOCOMPETENTE

2
– nereceptivitatea organismului la orice agent străin din punct de vedere genetic, inclusiv la mi/o şi toxinele lor. IMUNITATE – nereceptivitatea organismului la orice agent străin din punct de vedere genetic, inclusiv la mi/o şi toxinele lor. – capacitatea de apărare specifică a organismului faţă de agresori externi (virusuri, bacterii, fungi, protozoare, toxine) cât şi faţă de propriile molecule şi unele celule degradate sau modificate. IMUNITATE – capacitatea de apărare specifică a organismului faţă de agresori externi (virusuri, bacterii, fungi, protozoare, toxine) cât şi faţă de propriile molecule şi unele celule degradate sau modificate. Sarcina fundamentală a imunităţii – distincţia dintre moleculele şi celulele proprii (self) şi cele străine (non-self) Sarcina fundamentală a imunităţii – distincţia dintre moleculele şi celulele proprii (self) şi cele străine (non-self)

3
TIPURILE DE IMUNITATE I. Ereditară (naturală, de specie). Poate fi absolută (lipsa ţintei) sau relativă. II. Dobândită (achizitionata) 1. Activă - Naturală (postinfecţioasă) - Artificială (în urma vaccinării) 2. Pasivă - Naturală (transplacentară, prin laptele matern) - Naturală (transplacentară, prin laptele matern) - Artificială (administrarea Ac / seruri imune sau a - Artificială (administrarea Ac / seruri imune sau a limfocitelor) limfocitelor)
. Poate fi absolută (lipsa ţintei) sau relativă. II. Dobândită (achizitionata) 1. Activă - Naturală (postinfecţioasă) - Artificială (în urma vaccinării) 2. Pasivă - Naturală (transplacentară, pr")
4
Imunitatea dobândită se caracterizează prin: 1. Dezvoltare lenta si manifestare tardiva (câteva zile, săptămâni dupa contactul cu un antigen) 2. Specificitate faţă de antigen (capacitatea de a recunoaste si raspunde specific la numeroase substante straine, inclusiv agenti infectiosi) 3. Memorie imunologică (capacitatea de a elabora raspuns mai rapid, mai intens si eficace la intalniri repetate cu un antigen) 4. Lipsa reactivităţii (toleranţă)faţă de antigenele proprii Imunitatea dobândită poate fi: Imunitatea dobândită poate fi: I. Imunitate antibacteriană, antivirală, antimicotică, antitoxică, antitumorală, etc.
 2. Specificitate faţă de antigen (capacitatea de a recunoaste si raspunde specific la numeroase substante st")
5
II. II. În funcţie de mecanismele reacţiilor imune - Imunitate umorală, exercitată prin intermediul unor proteine numite anticorpi (Ac, Ig), produse de limfocitele B. Fiind secretate in sange si lichide biologice neutralizeaza si elimina microbii extracelulari si toxinele lor. - Imunitate celulară, eficienta in eliminarea parazitilor intracelulari sau a celulelor tumorale. Este exercitată prin intermediul limfocitelor T (citotoxicitate directa, activarea macrofagelor, celulelor NK, secreţia citokinelor) III. III. În dependenţă de persistenţa mi/o - Imunitate sterilă – se manifestă după eliminarea agenţilor patogeni din organism (ex.:rujeolă) - Imunitate nesterilă – nereceptivitatea se păstrează doar în perioada aflării mi/o în organism (tuberculoză, sifilis).
, produse de limfocitele B. Fiind secretate in sange si lichide biologice neutralizeaza si elimina microbii ext")
6
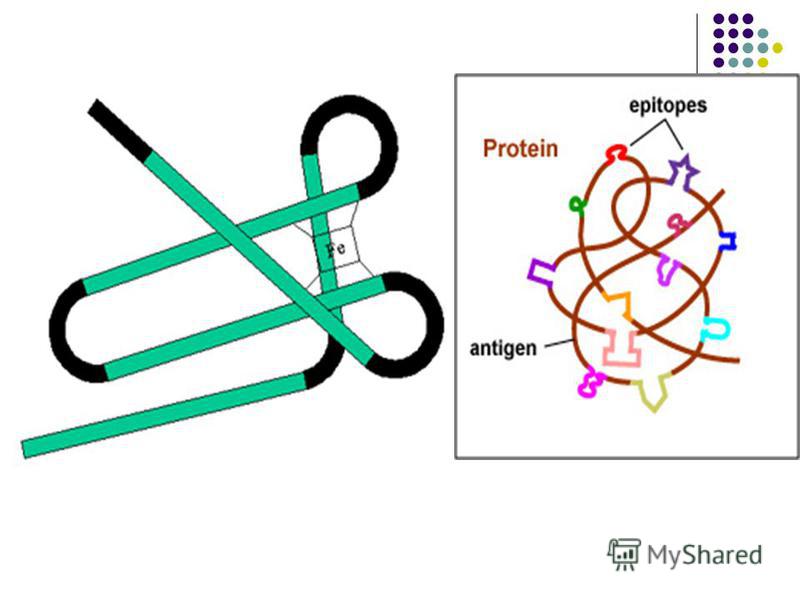
ANTIGENE – substanţe străine (non-self) de natură endo- sau exogenă capabile să declanşeze un răspuns imun (umoral, celular, toleranţă imunologică, memorie imunologică, paralizie imunologică). Proprietăţile de bază ale Ag: 1. Imunogenitatea – capacitatea Ag de a fi recunoscut ca străin şi de a induce răspuns imun specific 2. Antigenitatea (specificitatea) – capacitatea Ag de a interacţiona specific cu Ac sau cu receptorul pentru Ag complementar al limfocitelor sensibilizate
 de natură endo- sau exogenă capabile să declanşeze un răspuns imun (umoral, celular, toleranţă imunologică, memorie imunologică, paralizie imunologică). Proprietăţile de bază ale Ag: 1. Imunogenitatea – capacit")
7
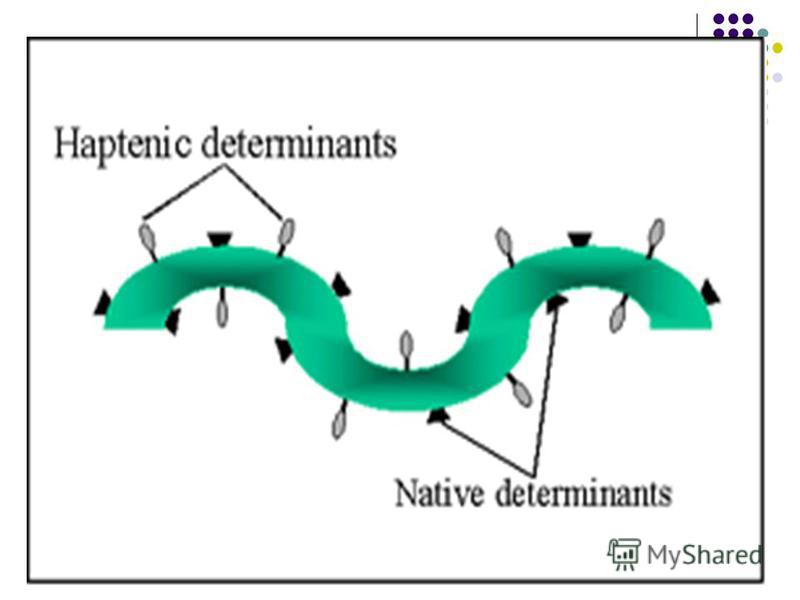
Antigenele care posedă ambele caractere sunt Ag complete. Antigenele care posedă ambele caractere sunt Ag complete. Cerinţele faţă de Ag complete: - Să fie substanţe străine - Să fie formate din gruparea carrier (purtător) şi epitopi (determinante antigenice) - Să aibă o greutate moleculară de peste 10 kDa - Să aibă structură chimică complexă (proteine, polizaharide, LPZ, etc) - Să aibă o configuraţie spaţială stabilă Antigenele incomplete (haptenele) posedă antigenitate dar sunt lipsite de imunogenitate. Antigenele incomplete (haptenele) posedă antigenitate dar sunt lipsite de imunogenitate.
 şi epitopi (determinante an")
8
Antigenele incomplete (haptenele): - Au masă moleculară mică - Pot deveni imunogene combinându-se cu macromolecule purtătoare (proteine sau polizaharide) Exemple de haptene: săruri ale metalelor grele (Cr, Ni), substanţe de origine vegetală, medicamente, coloranţi, oligonucleotide. – molecule proteice particulare (enterotoxinele stafilococice, toxina şocului toxic stafilococic, toxina exfoliativă a stafilococilor, nucleocapsida virusului rabic), capabile să stimuleze un număr mare de limfocite T (10- 40%). N – 0.01%. SuperAg provoacă reacţii imunopatologice. Superantigene – molecule proteice particulare (enterotoxinele stafilococice, toxina şocului toxic stafilococic, toxina exfoliativă a stafilococilor, nucleocapsida virusului rabic), capabile să stimuleze un număr mare de limfocite T (10- 40%). N – 0.01%. SuperAg provoacă reacţii imunopatologice.
: - Au masă moleculară mică - Pot deveni imunogene combinându-se cu macromolecule purtătoare (proteine sau polizaharide) Exemple de haptene: săruri ale metalelor grele (Cr, Ni), substanţe de origine vegetală, medicame")
10
Se disting arbitrar: - Ag solubile: proteine plasmatice, toxine, enzime, hormoni, etc - Ag figurate (celulare, corpusculare): celule, bacterii, paraziţi, etc. În funcţie de provenienţă se deosebesc: - Ag heterofile – Ag comune mai multor specii animale. Ex.: Ag polizaharidice Forssman, prezente în hematiile de cal, câine, oaie, cobai; sistemul Rh al eritrocitelor se întâlneşte la om şi maimuţele Macaccus rhesus, etc
: celule, bacterii, paraziţi, etc. În funcţie de provenienţă se deosebesc: - Ag heterofile – Ag comune mai multor specii anima")
11
- Ag heteroloage (xeno-Ag, hetero-Ag) – Ag provenite din organismul altei specii - Izo-Ag (alo-Ag) – Ag de grup în cadrul unei specii (sistemul ABO şi Rh, clasele de Ig) - Idioantigene – antigenele specifice unui individ. Corespund moleculelor CMH - Autoantigene – Ag proprii unui organism, devenite imunogene în anumite condiţii (spermatozoizii, tiroglobulina, insulina, cristalinul, ţesutul nervos, etc) - Exoantigene (provenite din compartimentul extracelular - bacterii, fungi, protozoare fagocitate) - Endoantigene (provenite din citoplasma celulelor, reprezintă proteine proprii modificate sau proteine virale sintetizate de celulele macroorganismului)
 – Ag provenite din organismul altei specii - Izo-Ag (alo-Ag) – Ag de grup în cadrul unei specii (sistemul ABO şi Rh, clasele de Ig) - Idioantigene – antigenele specifice unui individ. Corespund moleculelor CMH -")
12
Structura antigenică a celulei bacteriene Structura antigenică a celulei bacteriene I. Ag structurale (Ag O/R – somatice, LPZ, termostabile; Ag H – flagelar, proteic, termolabil; Ag K – capsular, termovariabil; Ag F – fimbrial) II. Ag solubile (enzime de patogenitate, exotoxine) proteine/GP de invelis, nucleoproteine, enzime Antigene virale: proteine/GP de invelis, nucleoproteine, enzime
 II. Ag solubile (enzim")
13
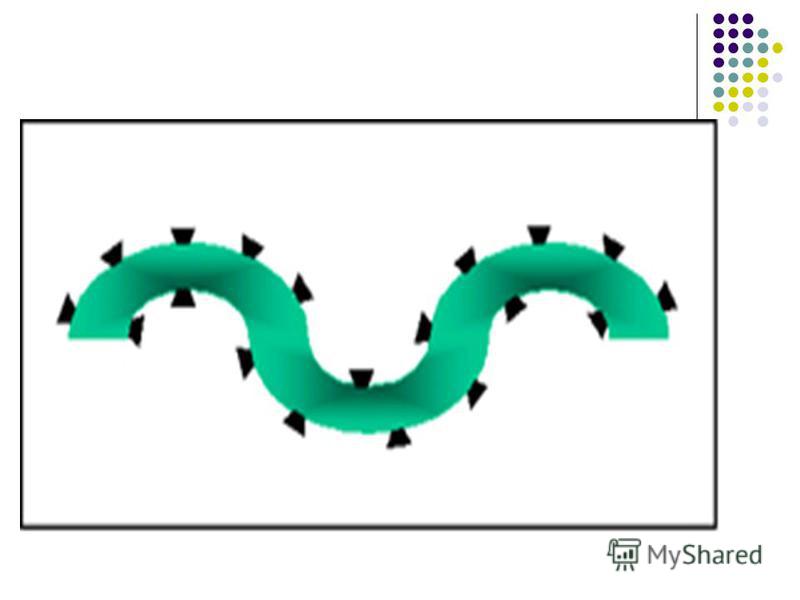
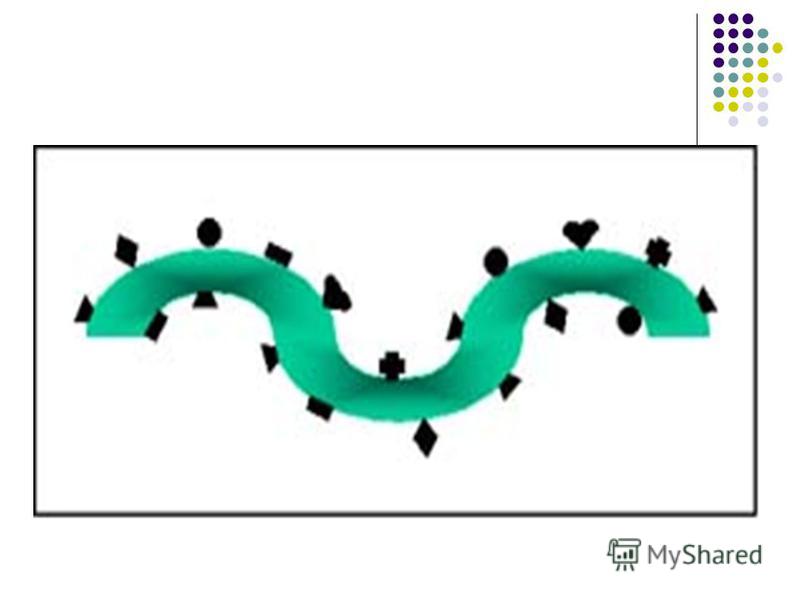
Determinantele antigenice (epitopii) Imunogenitatea este o caracteristică a întregii macromolecule Ag. Antigenitatea (specificitatea) este determinată de anumite secvenţe ale Ag. Suprafeţe limitate din macromolecula Ag apte să se combine cu Ac specifici sau cu receptorii de pe limfocitele sensibilizate se numesc determinante antigenice sau epitopi.
 Imunogenitatea este o caracteristică a întregii macromolecule Ag. Antigenitatea (specificitatea) este determinată de anumite secvenţe ale Ag. Suprafeţe limitate din macromolecula Ag apte să se combine cu Ac specif")
14
La glucide epitopul este format din 4-6 monozaharide. Polizaharidele sunt formate dintr-un epitop repetat sau dintr-un număr redus de epitopi diferiţi. La glucide epitopul este format din 4-6 monozaharide. Polizaharidele sunt formate dintr-un epitop repetat sau dintr-un număr redus de epitopi diferiţi. Polizaharidele sunt Ag T-independente, pot induce sinteza Ac fără intervenţia TL. Polizaharidele sunt Ag T-independente, pot induce sinteza Ac fără intervenţia TL.

15
Epitopul proteic este constituit din câţiva aminoacizi (AA). Epitopul proteic este constituit din câţiva aminoacizi (AA). - Epitopii liniari (secvenţiali) sunt determinaţi de structura primară a AA (8-30 AA) - Epitopii conformaţionali sunt determinaţi de structura secundară sau terţiară a moleculei proteice (juxtapoziţia în spaţiu a AA situaţi la distanţă). Se modifică la denaturarea proteinei.
. Epitopul proteic este constituit din câţiva aminoacizi (AA). - Epitopii liniari (secvenţiali) sunt determinaţi de structura primară a AA (8-30 AA) - Epitopii conformaţionali sunt determinaţ")
17
Fiecare moleculă proteică reprezintă un mozaic de epitopi, fie diferiţi, fie identici. Fiecare moleculă proteică reprezintă un mozaic de epitopi, fie diferiţi, fie identici. Proteinele sunt Ag T-dependente, deoarece ele induc sinteza Ac doar prin cooperarea dintre T şi B limfocite. Proteinele sunt Ag T-dependente, deoarece ele induc sinteza Ac doar prin cooperarea dintre T şi B limfocite. Numărul de epitopi de pe o moleculă imunogenă reprezintă valenţa Ag. Numărul de epitopi de pe o moleculă imunogenă reprezintă valenţa Ag.

21
Sistemul Imun reprezinta un ansamblu de celule, molecule şi tesuturi (organe) distribuite în tot organismul, care participa la instaurarea imunitatii. Functiile Sistemului Imun: 1. Prevenirea infectiilor şi eradicarea infecţiilor declanşate 2. Recunoasterea tesuturilor si proteinelor straine si raspunsul la ele 3. Protectie contra tumorilor
 distribuite în tot organismul, care participa la instaurarea imunitatii. Functiile Sistemului Imun: 1. Prevenirea infectiilor şi eradicarea infecţiilor declanşate 2. Recuno")
22
TESUTURILE SISTEMULUI IMUN Organele centrale (primare) ale SI – măduva osoasă şi timusul la vertebrate (ficatul în perioada embrionară), bursa Fabricius la păsări. bursa Fabricius la păsări. Apar primele în timpul vieţii Apar primele în timpul vieţiiembrionare. Rolul – sursa celulelor-stem, instruirea, maturizarea şi selectia celulelor imunocompetente (limfocitele T şi B).
 ale SI – măduva osoasă şi timusul la vertebrate (ficatul în perioada embrionară), bursa Fabricius la păsări. bursa Fabricius la păsări. Apar primele în timpul vieţii Apar primele în timpul vieţii")
23
În măduva osoasă se află celulele-stem, precursori ai T şi B-limfocitelor. Pre-T limfocitele migrează ulterior în timus, unde va avea loc instruirea şi maturizarea lor, devenind celule imunocompetente, capabile să recunoască specific un singur Ag (prin achiziţionarea unor receptori specifici). În măduva osoasă se află celulele-stem, precursori ai T şi B-limfocitelor. Pre-T limfocitele migrează ulterior în timus, unde va avea loc instruirea şi maturizarea lor, devenind celule imunocompetente, capabile să recunoască specific un singur Ag (prin achiziţionarea unor receptori specifici). Instruirea si maturizarea B-limfocitelor are loc în măduva osoasă. Aceste procese sunt independente de orice stimulare antigenică a organismului. Aceste procese sunt independente de orice stimulare antigenică a organismului. După părăsirea organelor centrale limfocitele nu mai revin aici, ele se stabilesc în organele limfoide secundare, recirculând prin sânge, limfă. După părăsirea organelor centrale limfocitele nu mai revin aici, ele se stabilesc în organele limfoide secundare, recirculând prin sânge, limfă.

24
Organele periferice (secundare) ale SI Organele periferice (secundare) ale SI În ele se realizează contactul dintre antigen, celulele prezentatoare de antigen ( CPA) şi celulele imuno-competente, cu inducerea unui raspuns imun. - Ganglionii limfatici (raspuns imun contra Ag transportate cu limfa) - Splina (raspuns imun contra Ag transportate cu sangele) - Formaţiunile limfoide ale mucoaselor digestive şi respiratorii (amigdale, plăci Peyer, apendice), ţesutul limfoid asociat tegumentului (raspuns imun contra Ag ce penetreaza prin epiteliu)
 ale SI Organele periferice (secundare) ale SI În ele se realizează contactul dintre antigen, celulele prezentatoare de antigen ( CPA) şi celulele imuno-competente, cu inducerea unui raspuns imun. - Ganglionii limfatici")
25
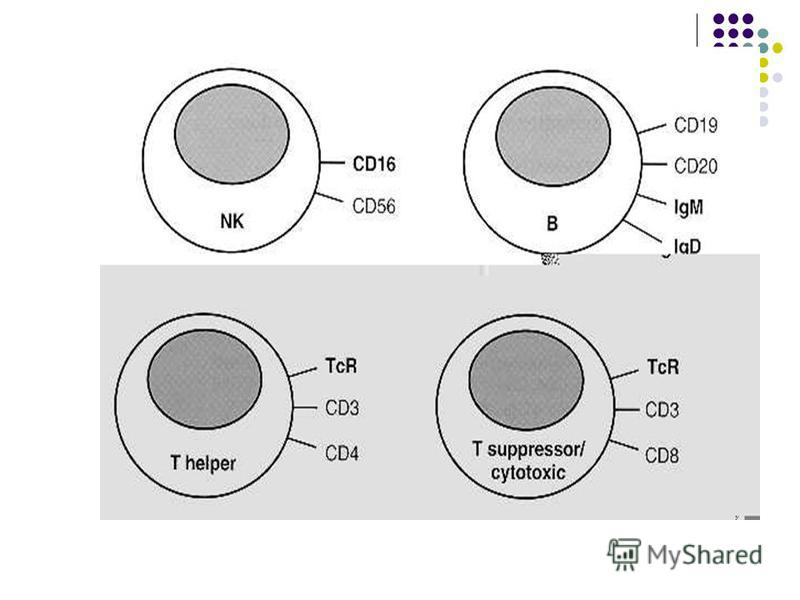
CELULELE SISTEMULUI IMUN 1. Limfocitele (celule imunocompetente, recunoasterea Ag) 2. Celulele prezentatoare de Ag (CPA) 2. Celulele prezentatoare de Ag (CPA) (captarea si prezentarea Ag microbiene ) 3. Celulele efectoare (eliminarea microbilor) LIMFOCITELE LIMFOCITELE Toate limfocitele provin din celule-stem ale maduvei osoase, cu o etapa ulterioara de maturizare si selectie. Limfocitele T si B sunt unicele celule ce poarta receptori specifici de Ag (celule imuno- competente), fiind mediatorii principali ai imunitatii dobandite
 2. Celulele prezentatoare de Ag (CPA) 2. Celulele prezentatoare de Ag (CPA) (captarea si prezentarea Ag microbiene ) 3. Celulele efectoare (eliminarea microbilor) LIMF")
26
Maturizarea limfocitelor B are loc în măduva osoasă prin intermediul interacţiunii directe cu celulele stromei, iar în stadiile tardive sub acţiunea citokinelor secretate de celulele stromale (în special IL-7). LB sunt supuse unei selectii pozitive in favoarea expresiei de receptori intacti si unei selectii negative contra recunoasterii puternice a antigenelor proprii. LB sunt supuse unei selectii pozitive in favoarea expresiei de receptori intacti si unei selectii negative contra recunoasterii puternice a antigenelor proprii. In organelor limfoide secundare limfocitele B se localizeaza in foliculii cortexului extern din ganglionii limfatici, foliculii din plãcile Peyer şi foliculii zonei periferice din pulpa albã a splinei (arii timo-independente). In organelor limfoide secundare limfocitele B se localizeaza in foliculii cortexului extern din ganglionii limfatici, foliculii din plãcile Peyer şi foliculii zonei periferice din pulpa albã a splinei (arii timo-independente).
. LB sunt supuse unei selectii pozitive i")
27
LB mature se caracterizează prin prezenţa următorilor receptori de suprafaţă: - Receptorul pentru Ag (BCR). Este reprezentat de molecule de imunoglobuline – monomeri de Ig M şi Ig D (forma membranara de anticorpi), asociate cu doua proteine Igα si Igβ. Interacţionează cu molecule antigenice din soluţii sau fixate pe membrane celulare (macromolecule native – proteine, lipide, glucide, acizi nucleici, grupari chimice mici) grupari chimice mici)
. Este reprezentat de molecule de imunoglobuline – monomeri de Ig M şi Ig D (forma membranara de anticorpi), asociate cu doua proteine Igα si Ig")
28
- Receptorul pentru fragmentul Fc al IgG - Receptor pentru fracţiunea C3d a complementului (CR2 sau CD21) - Receptori pentru interleukine (IL) - Proteine ale Complexului Major de Histocompatibilitate - CMH - Alte proteine specifice – CD79, CD19 – asociate BCR
 - Receptori pentru interleukine (IL) - Proteine ale Complexului Major de Histocompatibilitate - CMH - Alte proteine specifice – CD79, CD19 – aso")
29
Organismul contine clone diferite de BL (limfocite naive), fiecare cu BCR unic. BCR recunoaşte antigenele după configuraţia lor. BL stimulate de Ag se multiplică şi se diferenţiază în celule efectoare - plasmocite secretoare de Ig (Ac) şi în celule B-memorie.
, fiecare cu BCR unic. BCR recunoaşte antigenele după configuraţia lor. BL stimulate de Ag se multiplică şi se diferenţiază în celule efectoare - plasmocite secretoare de Ig (Ac) şi")
31
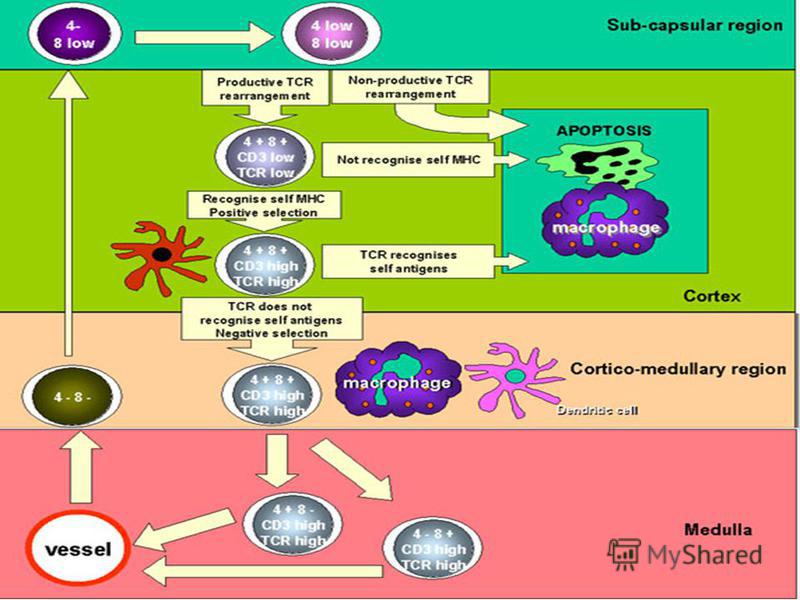
Precursorii limfocitelor T migrează în timus, unde sub influenţa celulelor stromale (IL-7) şi a corpusculilor Hassal se diferenţiază în limfocite T mature. Migraţia TL din cortex spre medulara timică este însoţită de achiziţia unor proteine de suprafaţă specifice (receptori: TCR, CD3, CD4, CD8, etc). La o etapa precoce de selectie doar limfocitele cu TCR functional vor supravietui. Dezvoltarea ulterioara a TL este caracterizată printr- o selecţie pozitivă (supravieţuiesc TL care recunosc moleculele Complexului Major de Histocompatibilitate (CMH) propriu). Aceste molecule sunt prezente pe suprafata celulelor dendritice si a macrofagelor din timus.
 şi a corpusculilor Hassal se diferenţiază în limfocite T mature. Migraţia TL din cortex spre medulara timică este însoţită de achiziţia unor proteine de supraf")
32
Limfocitele T care au trecut selecţia pozitivă, dar care sunt capabile să recunoască cu afinitate inalta peptide proprii asociate cu CMH suportă o selecţie negativă (moarte programată prin apoptoză). Selecţia constã în eliminarea clonelor de limfocite potenţial autoreactive (potenţial reactive faţã de moleculele proprii). Timusul selecţionează utilul, ignoră inutilul şi distruge nocivul. (Von Boehmer ). Timusul selecţionează utilul, ignoră inutilul şi distruge nocivul. ( Von Boehmer ). La etapa finala limfocitele T care sunt capabile sa recunoasca peptide in asociatie cu moleculele CMH I pierd receptorul CD4 (devenind celule TCD8), iar cele ce recunosc complexe peptid- CMH II pierd receptorul CD8 (devenind celule TCD4)
. Selecţia constã în eliminarea clonelor de limfocit")
34
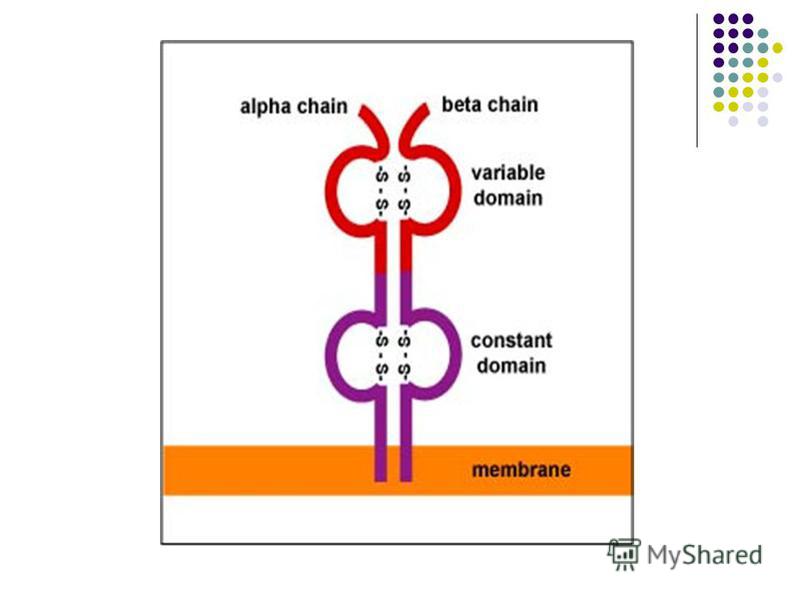
Receptorii TL: - TCR, r format din 2 catene polipeptidice α şi β (90%) sau γ şi δ. TCR poate recunoaşte doar peptide antigenice asociate cu molecule ale CMH, localizate pe suprafaţa unei Celule Prezentatoare de Antigen - CPA. Nu interacţionează cu Ag solubile. - TCR, receptorul pentru Ag - format din 2 catene polipeptidice α şi β (90%) sau γ şi δ. TCR poate recunoaşte doar peptide antigenice asociate cu molecule ale CMH, localizate pe suprafaţa unei Celule Prezentatoare de Antigen - CPA. Nu interacţionează cu Ag solubile. TCR recunoaste simultan peptidul antigenic si unele reziduri ale moleculei CMH. - CD3 (Cluster Differentiation 3) – asociat TCR, ajută la transmiterea în celulă a semnalului de recunoaştere a Ag si de activare a limfocitului - CD2– prezent la toate TL, fixează hematii, poate fi depistat prin testul de formare a rozetelor.
 sau γ şi δ. TCR poate recunoaşte doar peptide antigenice asociate cu molecule ale CMH, localizate pe suprafaţa unei Celule Prezentatoare de Antigen - CPA. Nu interacţionează cu Ag")
36
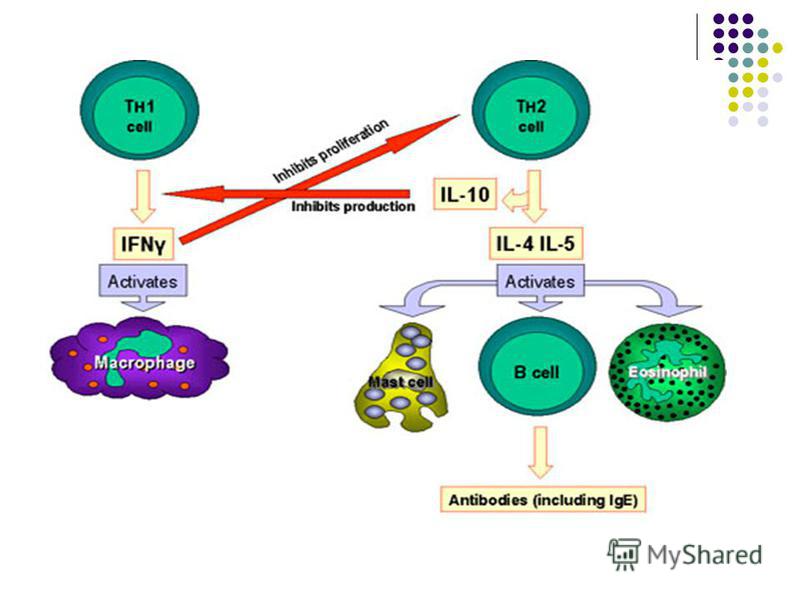
-Unii markeri (receptori) de suprafaţă definesc 2 varietăţi principale de LT: limfocitele T CD4+ şi TCD8+ - CD4 – exprimat pe 60% din TL. Recunosc antigenele peptidice asociate cu moleculele CMH clasa II (exprimate pe celule dendritice, macrofage şi LB). - CD4 – exprimat pe 60% din TL. Recunosc antigenele peptidice asociate cu moleculele CMH clasa II (exprimate pe celule dendritice, macrofage şi LB). Limfocitele TCD4 (T4) au funcţia de coordonator central al răspunsului imun (participa in stimularea raspunsul imun umoral si celular) şi la activare se diferenţiază în subpopulaţia de Thelper (Th, limfocite T auxiliare). Limfocitele TCD4 (T4) au funcţia de coordonator central al răspunsului imun (participa in stimularea raspunsul imun umoral si celular) şi la activare se diferenţiază în subpopulaţia de Thelper (Th, limfocite T auxiliare). Th acţionează prin producerea citokinelor. În funcţie de profilul de citokine elaborate se disting 2 subpopulatii/clone de Th: Th1 şi Th2.
 de suprafaţă definesc 2 varietăţi principale de LT: limfocitele T CD4+ şi TCD8+ - CD4 – exprimat pe 60% din TL. Recunosc antigenele peptidice asociate cu moleculele CMH clasa II (exprimate pe celule dendritice, macrofage şi")
37
Diferenţierea în Th1 este favorizată de IL-12 produsă de macrofage si celule dendritice. Citokinele secretate de Th1 (IFN-γ, IL-2, TNF) stimulează răspunsul imun celular şi hipersensibilitatea tardivă (activarea macrofagelor si stimularea functiilor microbicide ale fagocitelor, stimularea proliferarii şi diferenţierii limfocitelor T citotoxice (Tc), activarea procesului inflamator). De asemenea Th1 stimulează producerea Ac opsonizanţi si fixatori de complement – IgG1, IgG3 (anticorpi ce favorizeaza fagocitoza)
 stimulează răspunsul imun celular şi hipersensibilitatea tardivă (activarea macrofagelor si stimularea functiilor")
39
Th2 recunosc Ag prezentate în special de B- limfocite. Secretă IL-4, IL-5, IL-10 şi IL-13. IL-4 stimulează producerea Ac Ig E, iar IL-5 activează eozinofilele (rol in reactiile de hipersensibilitate imediate/anafilactice si in eliminarea helmintilor). De asemenea Th2 determina sinteza anticorpilor neutralizanti IgG. Există o competiţie între cele două tipuri de Th: IFN-γ inhibă diferenţierea şi proliferarea Th2, iar IL-4, IL-10 şi IL-13 inhibă activitatea Th1.
")
42
Tfh – stimulează diferenţierea BL în plasmocite cu sinteza Ig din toate clasele cu excepţia Ig E. Produc IL-21.Se intâlnesc in foliculii din ganglionii limfatici. Th17 – asigură protecţia suprafeţelor (tegumenmt, mucoase) contra bacteriilor extracelulare. Produc IL-17 şi IL-22). Treg – imunosupresie, acţiune antiinflamatoare.
 contra bacteriilor extracelu")
43
- CD8 – exprimat pe 40% LT. Recunosc antigenele peptidice asociate cu moleculele CMH clasa I (exprimate pe toate celulele nucleate). Pot produce citokine, dar în special manifestă activitate citotoxica. La activare se diferenţiază în limfocite T citotoxice (Tc) şi T- reglatoare (Treg). Tc distrug celulele infectate cu virus, cu bacterii intracelulare sau celulele canceroase, intervin în respingerea grefelor. Mecanisme: secretia porinelor, inducerea fragmentarii ADN-ului si a mortii prin apoptoza a celulelor. Mecanisme: secretia porinelor, inducerea fragmentarii ADN-ului si a mortii prin apoptoza a celulelor. T-memorie asigură memoria imunologică
. Pot produce citokine, dar în special manifestă activitate citotoxica. La activare se diferenţiază în limfocite T cito")
45
Limfocitele B şi T naive sunt celulele care inca nu s-au întâlnit cu un Ag. Au functia de recunoastere a Ag. Populeaza organele limfoide periferice si recircula prin ele in cautarea Ag specifice. După interacţiunea cu Ag urmeaza activarea lor, proliferarea şi diferenţierea în celule efectoare (LB – în plasmocite producatoare de Ac, LT în Th, Tc). Celulele (Th, Tc) si moleculele (Ac) efectoare migreaza spre locul infectiei unde vor elimina microbii sau neutraliza toxinele lor prin diferite mecanisme. Celulele efectoare au o durata de viata scurta si mor prin apoptoza dupa eliminarea Ag. Celulele efectoare au o durata de viata scurta si mor prin apoptoza dupa eliminarea Ag. LB reprezintă 10-15% din populaţia limfocitară, cu o durată de viaţă scurtă, de 3-5 zile, iar LT constituie 70% cu o durată de viaţă lungă (luni, ani). Raportul limfocitelor CD4/CD8 este de 1,5

46
Alte celule limfocitare (nule, neimuno- competente) – 15% - Celule NK, capabile să distrugă spontan celule canceroase şi celule infectate cu virus. In raspuns la IL-12 produsa de macrofage celulele NK secreta IFN-γ, care activeaza macrofagele pentru a distruge microbii fagocitati. - Celule K, posedă pe membrana lor receptori pentru fragmentul Fc al Ig. Distrug celulele-ţintă acoperite cu Ac (Ig) – fenomen de citotoxicitate Ac -dependentă.
 – 15% - Celule NK, capabile să distrugă spontan celule canceroase şi celule infectate cu virus. In raspuns la IL-12 produsa de macrofage celulele NK secreta IFN-γ, care activeaza macrofagele pentru")
51
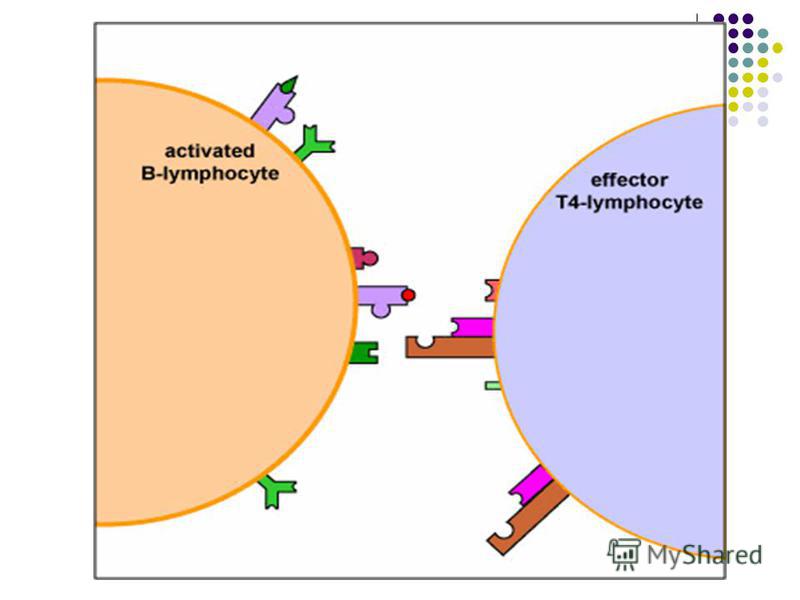
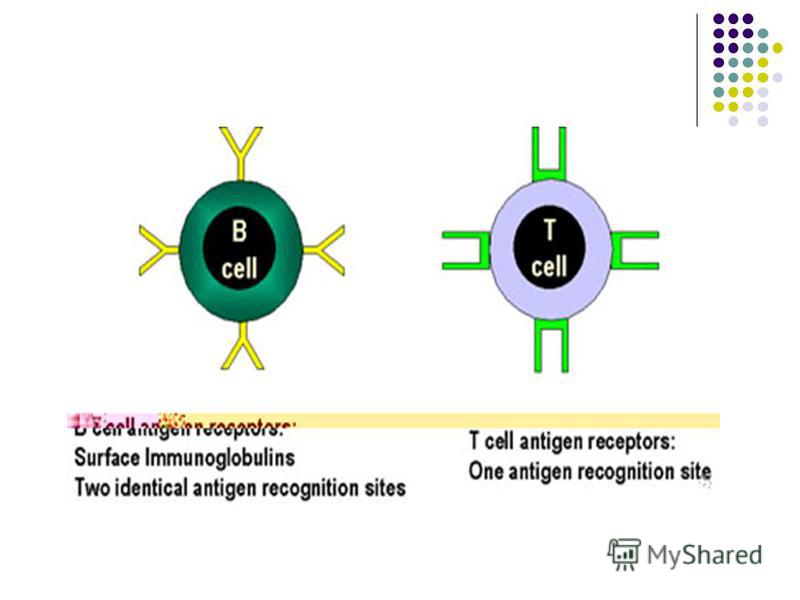
CELULELE PREZENTATOARE DE ANTIGEN (CPA) BL recunosc direct prin intermediul BCR epitopi antigenici conformaţionali solubili sau fixaţi pe membrane celulare. BL recunosc direct prin intermediul BCR epitopi antigenici conformaţionali solubili sau fixaţi pe membrane celulare. TL pot recunoaşte numai epitopi proteici liniari asociaţi cu molecule ale CMH (restricţie CMH) expuşi pe suprafaţa unor celule specializate – CPA TL pot recunoaşte numai epitopi proteici liniari asociaţi cu molecule ale CMH (restricţie CMH) expuşi pe suprafaţa unor celule specializate – CPA
 BL recunosc direct prin intermediul BCR epitopi antigenici conformaţionali solubili sau fixaţi pe membrane celulare. BL recunosc direct prin intermediul BCR epitopi antigenici conformaţionali solubili sau fixaţ")
52
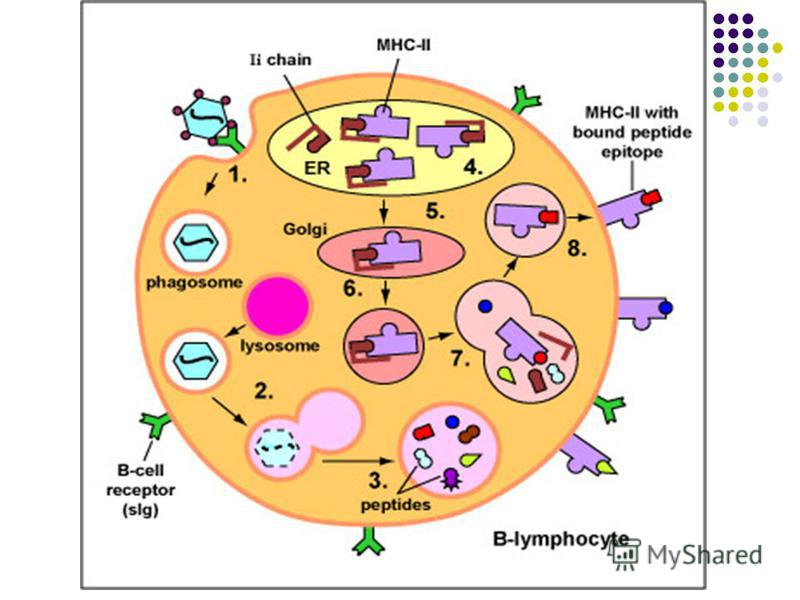
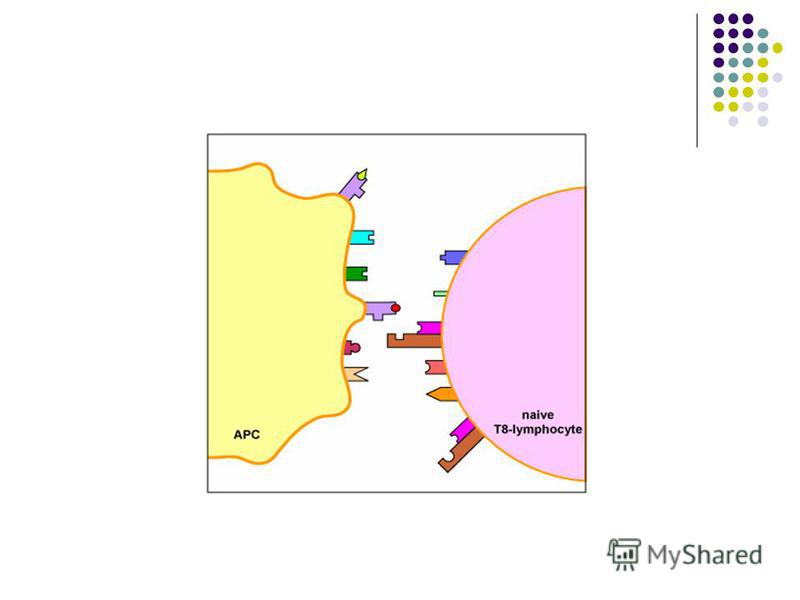
CPA profesioniste includ celulele dendritice foliculare şi tisulare, monocitele / macrofagele şi limfocitele B. Ele se intalnesc la posibilele porti de intrare ale agentilor patogeni. Funcţiile CPA 1. Captarea, fragmentarea si procesarea (processing-ul) Ag proteice cu expunerea pe suprafaţa sa a peptidelor antigenice (epitopi liniari) asociate cu moleculele CMH. 2. Transportarea acestor complexe spre tesutul limfoid periferic, unde complexul CMH-peptid va fi prezentat TL naiv cu TCR specific, declanşand activarea lui. CPA care exprima peptide asociate cu CMH I vor interactiona cu limfocite TCD8, iar CPA care exprima peptide in asociatie cu CMH II – cu TCD4 3. Producerea semnalelor secundare de activare a LT (secreţia unor citokine, expresia unor molecule de co- stimulare)

54
Toate celulele nucleate de asemenea sunt capabile sa prezinte peptide antigenice in asociatie cu moleculele CMH de clasa II (CPA neprofesioniste).
.")
55
COMPLEXUL MAJOR DE HISTOCOMPATIBILITATE Moleculele CMH reprezintă un ansamblu unic de glicoproteine exprimate pe suprafaţa celulelor organismului, care sunt veritabili markeri / antigene de identitate ai fiecărui individ. Rolul moleculelor CMH: - Participă la prezentarea antigenelor peptidice limfocitelor T - Determina acceptarea sau respingerea grefelor tisulare

56
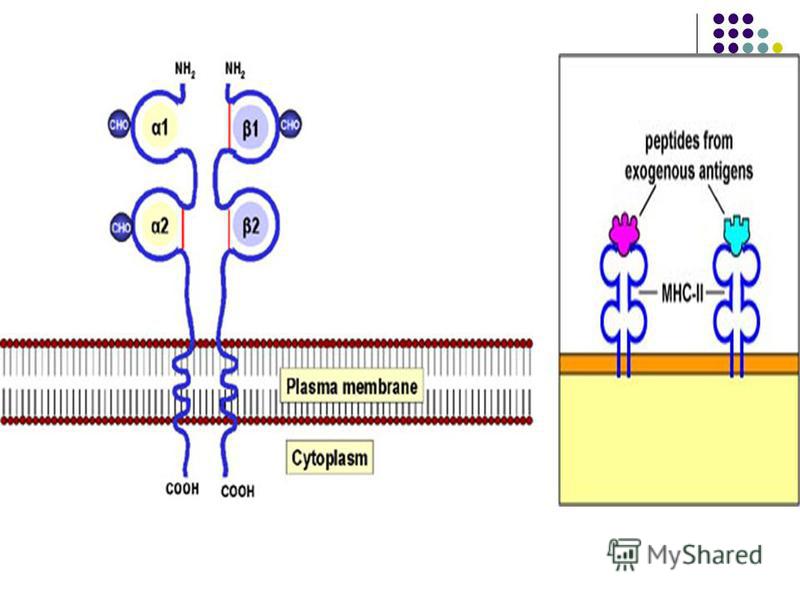
CMH este transmis genetic. Genele CMH sunt situate pe cromozomul 6 la om, locus numit HLA (Human Leukocyte Antigen). CMH este transmis genetic. Genele CMH sunt situate pe cromozomul 6 la om, locus numit HLA (Human Leukocyte Antigen). Moleculele CMH sunt formate din 2 catene polipeptidice, constituite din domenii extracelulare, un fragment transmembranar şi o regiune intracitoplasmatică. Structura tridimensională a moleculei duce la formarea unei cavităţi, la fundul căreia se află receptorii care vor interacţiona cu peptide antigenice proprii sau străine. Alţi receptori ai CMH interacţionează cu moleculele CD4 sau CD8 de pe TL. Moleculele CMH sunt formate din 2 catene polipeptidice, constituite din domenii extracelulare, un fragment transmembranar şi o regiune intracitoplasmatică. Structura tridimensională a moleculei duce la formarea unei cavităţi, la fundul căreia se află receptorii care vor interacţiona cu peptide antigenice proprii sau străine. Alţi receptori ai CMH interacţionează cu moleculele CD4 sau CD8 de pe TL.
. CMH este transmis genetic. Genele CMH sunt situate pe cromozomul 6 la om, locus numit HLA (Human Leukocyte Antigen). Moleculele CMH s")
58
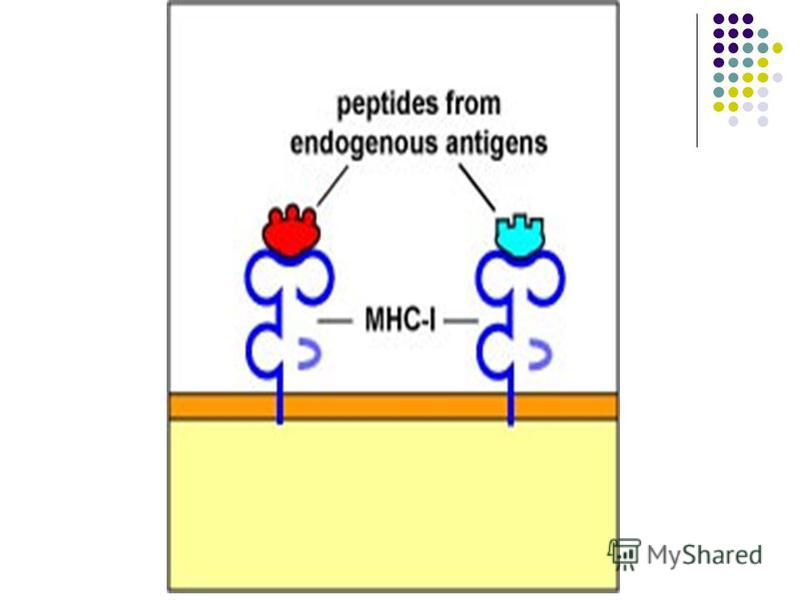
Se disting molecule (antigene) CMH de clasa I şi de clasa II, sintetizate in reticulul endoplasmatic celular. Se disting molecule (antigene) CMH de clasa I şi de clasa II, sintetizate in reticulul endoplasmatic celular. Moleculele CMH de clasa I sunt exprimate pe toate celulele nucleate ale organismului (inclusiv CPA). CMH I participă la prezentarea peptidelor antigenice scurte (9 AA) provenite din proteine antigenice citoplasmatice produse de virusuri ce paraziteaza celula, de bacterii facultativ intracelulare care au reusit sa paraseasca fagosoma sau de gene celulare mutate sau alterate (ex. in cazul tumorilor). Peptidele sunt obţinute în cursul degradării in citoplasma (de către proteasome) a proteinelor mentionate mai sus (antigene endogene). CMH I recunoaste si interacţionează cu receptorul CD8 de pe T-limfocite.
 CMH de clasa I şi de clasa II, sintetizate in reticulul endoplasmatic celular. Se disting molecule (antigene) CMH de clasa I şi de clasa II, sintetizate in reticulul endoplasmatic celular. Moleculele CMH de clasa I sunt")
61
- Complexul peptid/CMH I de pe suprafaţa Celulelor Prezentatoare de Antigen profesioniste (macrofage, celule dendritice) poate fi recunoscut de complexul TCR/CD8 complementar de pe limfocite T CD8 naive (rezultă activarea limfocitelor, diferenţierea lor în limfocite Tc - iniţierea răspunsului imun celular)
 poate fi recunoscut de complexul TCR/CD8 complementar de pe limfocite T CD8 naive (rezultă activarea limfocitelor, diferenţierea")
63
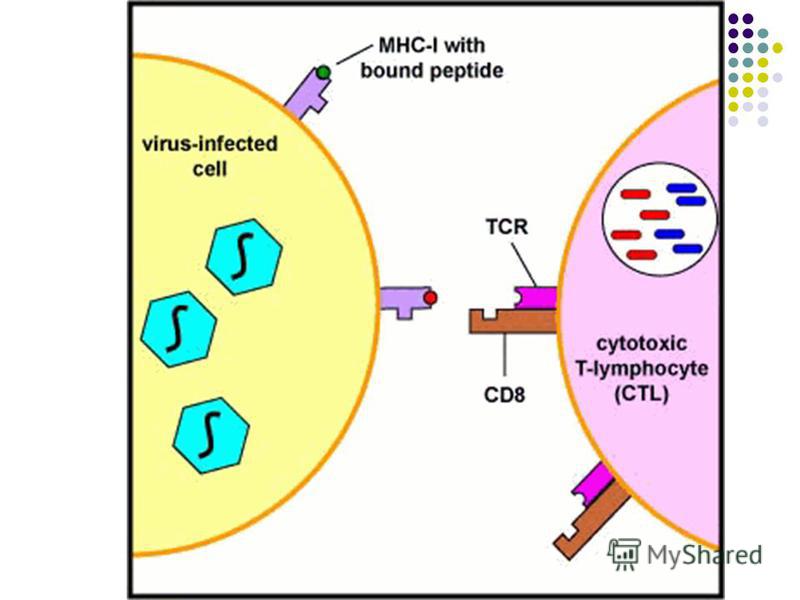
- Complexul peptid /CMH I de pe suprafaţa celulelor infectate sau a celulelor tumorale (CPA neprofesioniste) poate fi recunoscut de complexul TCR/CD8 complementar de pe limfocitele efectoare Tc (rezultă distrugerea celulelor ce conţin antigenul endogen)
 poate fi recunoscut de complexul TCR/CD8 complementar de pe limfocitele efectoare Tc (rezultă distrugerea celulelor ce conţin antigenul endog")
66
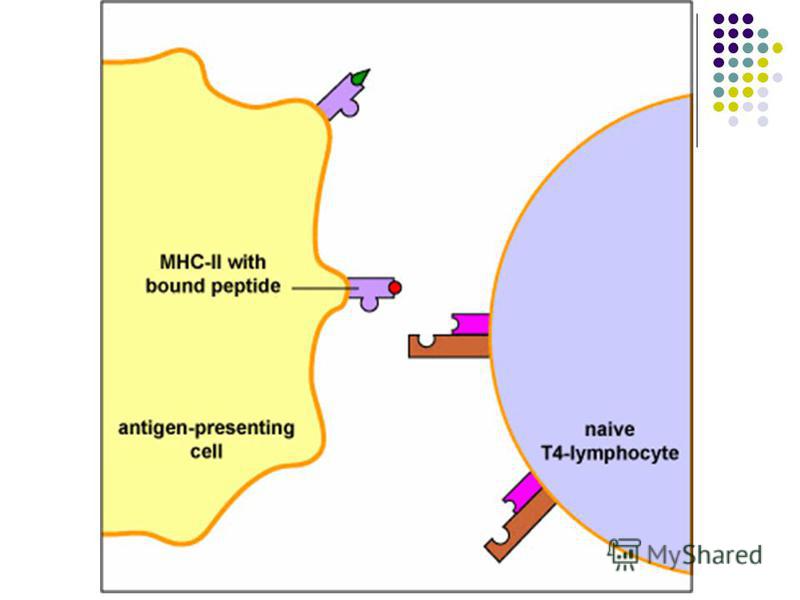
- Moleculele CMH de clasa II sunt exprimate pe CPA specializate în prezentarea Ag limfocitelor TCD4: limfocite B, celule dendritice, monocite, macrofage. Ele pot prezenta peptide din AA derivate din degradarea în fagolizosome a Ag exogene (bacterii, protozoare, fungi, virioni liberi). Complexul peptid/CMH II de pe celulele prezentatoare de Ag interacţionează cu complexul TCR/CD4 de pe limfocitele TCD4 naive sau efectoare (Th1, Th2). Astfel moleculele CMH II sunt implicate în ambele tipuri de imunitate: umorală şi celulară.

71
Prezentarea Ag limfocitelor T. Ag este prezentat receptorului TCR sub formă de peptide asociate moleculelor CMH de clasa I sau II de pe CPA. Prezentarea Ag limfocitelor T. Ag este prezentat receptorului TCR sub formă de peptide asociate moleculelor CMH de clasa I sau II de pe CPA. Limfocitele T CD8 recunosc în special Ag endogene, provenite din proteine citoplasmatice (molecule proprii modificate sau proteine virale) degradate în proteasome. Aceste peptide din 9 AA sunt prezentate în asociaţie cu moleculele CMH de clasa I de pe macrofage, celule dendritice, dar şi de pe suprafaţa tuturor celulelor nucleate ale organismului. Limfocitele T CD8 recunosc în special Ag endogene, provenite din proteine citoplasmatice (molecule proprii modificate sau proteine virale) degradate în proteasome. Aceste peptide din 9 AA sunt prezentate în asociaţie cu moleculele CMH de clasa I de pe macrofage, celule dendritice, dar şi de pe suprafaţa tuturor celulelor nucleate ale organismului.

74
Limfocitele T CD4 recunosc Ag exogene (provenite din bacterii, protozoare, virusuri libere). Prezentarea este asigurată de către celule dendritice tisulare, celule Langerhans din epidermă, limfocite B, monocite/macrofage (CPA profesioniste). Limfocitele T CD4 recunosc Ag exogene (provenite din bacterii, protozoare, virusuri libere). Prezentarea este asigurată de către celule dendritice tisulare, celule Langerhans din epidermă, limfocite B, monocite/macrofage (CPA profesioniste). După captarea Ag şi fragmentarea lor (în fagolizosomi) peptidele antigenice selectate ( AA) sunt ataşate de moleculele CMH II şi expuse pe suprafaţa CPA pentru a fi prezentate limfocitelor TCD4 naive, urmând activarea şi diferenţierea lor în celule efectori Th1 sau Th2 După captarea Ag şi fragmentarea lor (în fagolizosomi) peptidele antigenice selectate ( AA) sunt ataşate de moleculele CMH II şi expuse pe suprafaţa CPA pentru a fi prezentate limfocitelor TCD4 naive, urmând activarea şi diferenţierea lor în celule efectori Th1 sau Th2
. Prezentarea este asigurată de către celule dendritice tisulare, celule Langerhans din epidermă, limfocite B, monocite/macrofage (CPA profesioniste). Limfocit")
77
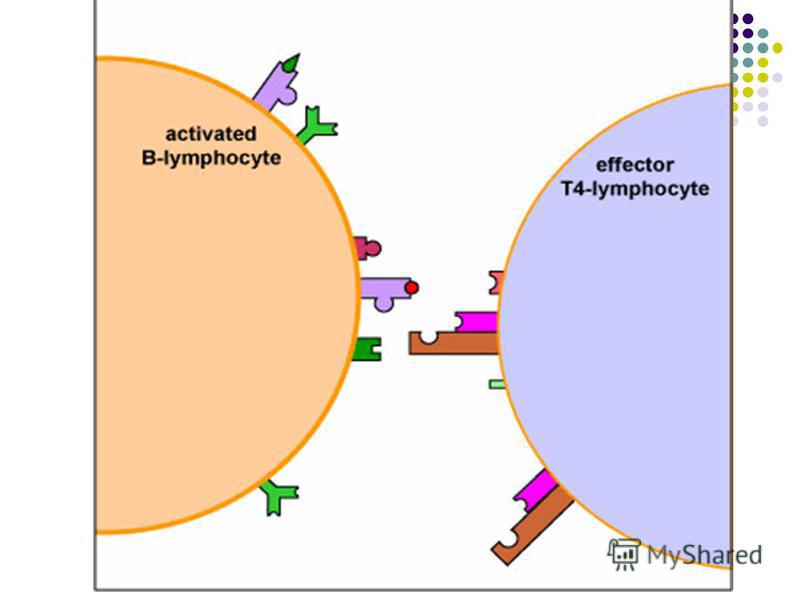
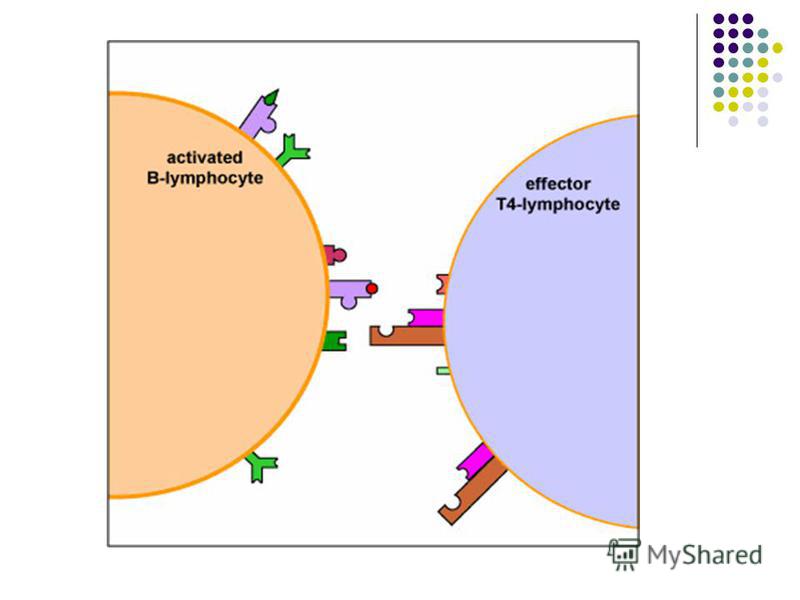
Prezentarea Ag limfocitelor B poate fi realizata prin intermediul celulelor dendritice foliculare din splină şi ganglioni limfatici. Aceste celule nefagocitare fixează prin intermediul receptorilor membranari Ag polizaharidice sau proteice prezentându-le ulterior limfocitelor B. Prezentarea Ag limfocitelor B poate fi realizata prin intermediul celulelor dendritice foliculare din splină şi ganglioni limfatici. Aceste celule nefagocitare fixează prin intermediul receptorilor membranari Ag polizaharidice sau proteice prezentându-le ulterior limfocitelor B.

Еще похожие презентации в нашем архиве:













 Ig prezintă glicoproteine din fracţia γ -globulinelor. Se disting Ig membranare (BCR) şi Ig.")
 Ig prezintă glicoproteine din fracţia γ - globulinelor. Se disting Ig membranare (BCR) şi Ig.")

 Starea solidă: au formă proprie; rigiditate; au volum propriu (incompresibilitate). b) Starea lichidă:")
 SEDIUL ACTIVITĂŢILOR METABOLICE SEDIUL ACTIVITĂŢILOR METABOLICE DIAMETRU : μ m mm DIAMETRU.")

")




, constă din: plasmă (partea lichidă) - 52-58% elemente.")







