Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
Кировская государственная медицинская академия Кафедра биологической химии Лекция: КОНЕЧНЫЕ ПРОДУКТЫ АЗОТИСТОГО ОБМЕНА. КОНЕЧНЫЕ ПРОДУКТЫ АЗОТИСТОГО ОБМЕНА. БИОСИНТЕЗ МОЧЕВИНЫ Зав. кафедрой: доктор медицинских наук, профессор Цапок Петр Иванович

2
План лекции 1. Конечные продукты азотистого обмена: соли аммония, мочевина и мочевая кислота. 1. Конечные продукты азотистого обмена: соли аммония, мочевина и мочевая кислота. 2. Обезвреживание аммиака: синтез глутамина и карбамилфосфата, восстановительное аминирование 2- оксоглутарата. 2. Обезвреживание аммиака: синтез глутамина и карбамилфосфата, восстановительное аминирование 2- оксоглутарата. 3. Глутамин как донор амидной группы при синтезе ряда соединений. Глутаминаза почек, образование и выведение солей аммония. Адаптивная активация глутаминазы почек при ацидозе. 3. Глутамин как донор амидной группы при синтезе ряда соединений. Глутаминаза почек, образование и выведение солей аммония. Адаптивная активация глутаминазы почек при ацидозе.

3
План лекции 4. Биосинтез мочевины. 4. Биосинтез мочевины. 5. Связь орнитинового цикла с превращениями фумаровой и аспарагиновой кислот; происхождение атомов азота мочевины. 5. Связь орнитинового цикла с превращениями фумаровой и аспарагиновой кислот; происхождение атомов азота мочевины. 6. Биосинтез мочевины как механизм предотвращения образования аммиака. Уремия. 6. Биосинтез мочевины как механизм предотвращения образования аммиака. Уремия.

4
КОНЕЧНЫЕ ПРОДУКТЫ : АММИАК КОНЕЧНЫЕ ПРОДУКТЫ : АММИАК Деградация аминокислот происходит преимущественно в печени. При этом непосредственно или косвенно освобождается аммиак. Значительные количества аммиака образуются при распаде пуринов и пирамидинов. Деградация аминокислот происходит преимущественно в печени. При этом непосредственно или косвенно освобождается аммиак. Значительные количества аммиака образуются при распаде пуринов и пирамидинов.

6
ТОКСИЧНОСТЬ АММИАКА Аммиак - NH 3 является клеточным ядом. При высоких концентрациях он повреждает главным образом нервные клетки (гепатаргическая кома). Аммиак - NH 3 является клеточным ядом. При высоких концентрациях он повреждает главным образом нервные клетки (гепатаргическая кома). В норме распад 70 г АК в сутки ведет к концентрации NH 3 в крови 60 мкмоль/л, что в 100 раз меньше концентрации глюкозы в крови. В норме распад 70 г АК в сутки ведет к концентрации NH 3 в крови 60 мкмоль/л, что в 100 раз меньше концентрации глюкозы в крови.
. Аммиак - NH 3 является клеточным ядом. При высоких концентрациях он повреждает главным образом н")
7
Токсичность аммиака В опытах на кроликах концентрация В опытах на кроликах концентрация NH 3 3 ммоль/л вызывала смерть! NH 3 3 ммоль/л вызывала смерть! Причины токсичности: Причины токсичности: 1. при рН крови в виде NH 4 +, проникает через плазм. и МХ мембраны с большим трудом. 1. при рН крови в виде NH 4 +, проникает через плазм. и МХ мембраны с большим трудом.

8
Нейтр. мол. своб. NH 3 легко проходят эти мембраны. При рН 7.4 только 1% NH 3 от общего количества аммиака проникает в клетки мозга и МХ. Нейтр. мол. своб. NH 3 легко проходят эти мембраны. При рН 7.4 только 1% NH 3 от общего количества аммиака проникает в клетки мозга и МХ.

9
Причины токсичности 2. NH 3 + а-КГ + НАДФН NH 3 + а-КГ + НАДФН 2 - Глу Н 2 О Глу + НАДФ + Н 2 О Отток альфа- КГ из фонда ЦТК и как следствие – снижение скорости окисления глюкозы

10
Токсичность аммиака Аммиак настолько токсичен, что должен быть немедленно удален посредством того или иного экскреторного механизма, либо путем включения в какое-то другое азотсодержащее соединение, не обладающее подобной токсичностью. Аммиак настолько токсичен, что должен быть немедленно удален посредством того или иного экскреторного механизма, либо путем включения в какое-то другое азотсодержащее соединение, не обладающее подобной токсичностью.

11
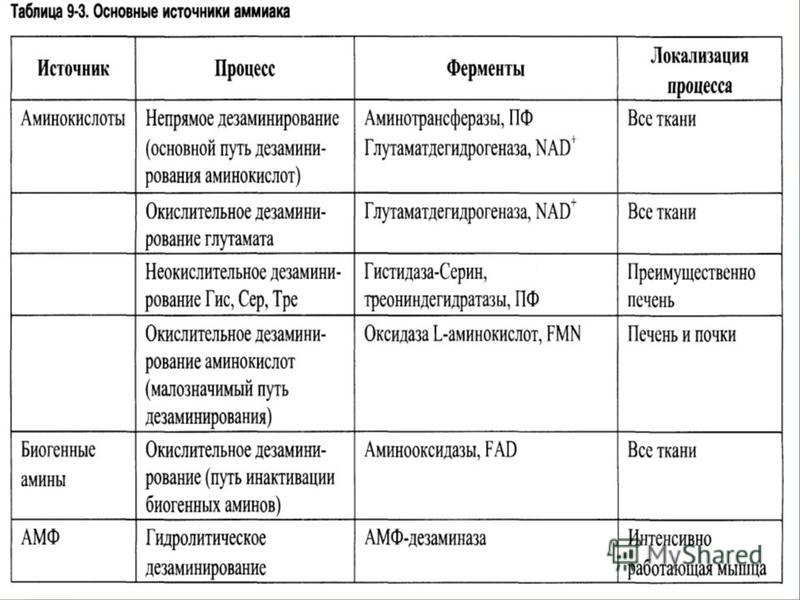
Механизмы детоксикации аммиака 1. Синтез глутамина: Глн, аспарагина: Асн. 1. Синтез глутамина: Глн, аспарагина: Асн. 2. Синтез мочевины. 2. Синтез мочевины. 3. Аминирование а-КГ --> Глу. 3. Аминирование а-КГ --> Глу. 4. Амидирование белков. 4. Амидирование белков.

12
Механизмы детоксикации аммиака 5. Синтез пурин. и пирамид. структур. 5. Синтез пурин. и пирамид. структур. 6. Нейтрализация в почках кислотами и выделение с мочой аммонийных солей. 6. Нейтрализация в почках кислотами и выделение с мочой аммонийных солей.

13
Обезвреживание аммиака В организмах автотрофов большая часть образующегося аммиака может вновь использоваться для синтеза новых клеточных структур. Гетеротрофы же обычно получают с пищей значительное количество белка, усвоение которого легко может привести к накоплению большого количества конечных продуктов азотистого обмена. Удаление этих отходов требует создания соответствующего аппарата. В организмах автотрофов большая часть образующегося аммиака может вновь использоваться для синтеза новых клеточных структур. Гетеротрофы же обычно получают с пищей значительное количество белка, усвоение которого легко может привести к накоплению большого количества конечных продуктов азотистого обмена. Удаление этих отходов требует создания соответствующего аппарата.

14
Обезвреживание аммиака Организм, живущий в водной среде, может выделять аммиак непосредственно, поскольку он будет немедленно разбавлен водой, не оказывая никакого или почти никакого вредного влияния на клетки. Экскреция аммиака у животных, обитающих в засушливых областях, потребовала бы для его разведения использования собственных водных ресурсов. Организм, живущий в водной среде, может выделять аммиак непосредственно, поскольку он будет немедленно разбавлен водой, не оказывая никакого или почти никакого вредного влияния на клетки. Экскреция аммиака у животных, обитающих в засушливых областях, потребовала бы для его разведения использования собственных водных ресурсов. Поэтому у многих видов аммиак превращается в организме в некоторые другие соединения, обладающие меньшей токсичностью. Поэтому у многих видов аммиак превращается в организме в некоторые другие соединения, обладающие меньшей токсичностью.

15
Восстановительное аминирование Большинство организмов обладает способностью реутилизировать аммиак за счет реакции, катализируемой глутаматдегидрогеназой. Большинство организмов обладает способностью реутилизировать аммиак за счет реакции, катализируемой глутаматдегидрогеназой. А-Кетоглутарат + NH3 + НАДФН.Н+ А-Кетоглутарат + NH3 + НАДФН.Н+ Глутамат + НАДФ+. Глутамат + НАДФ+. Это восстановительное аминирование. Это восстановительное аминирование. Однако все же некоторая часть образовавшегося аммиака остается неиспользованной и в конце концов выводится из организма беспозвоночных и позвоночных либо в свободном виде, либо в форме мочевой кислоты, либо в форме мочевины. Однако все же некоторая часть образовавшегося аммиака остается неиспользованной и в конце концов выводится из организма беспозвоночных и позвоночных либо в свободном виде, либо в форме мочевой кислоты, либо в форме мочевины.

16
Восстановительное аминирование

17
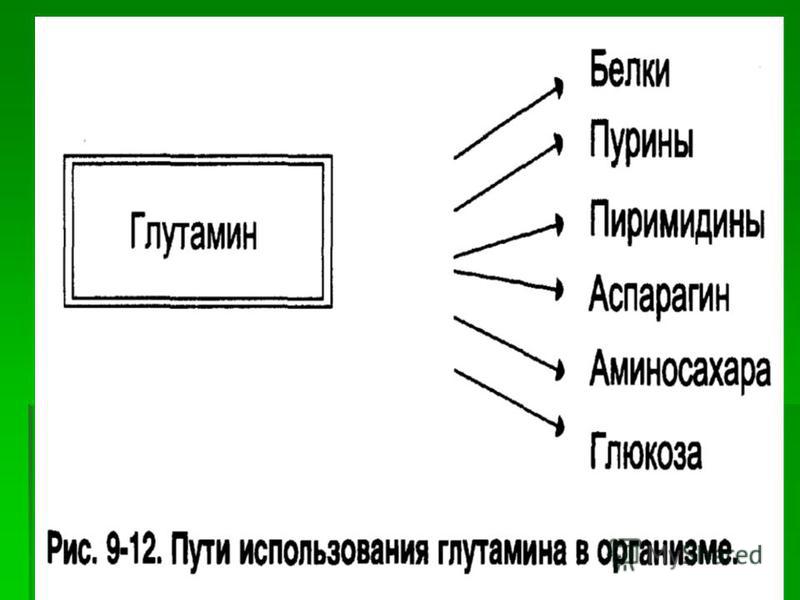
Биосинтез глутамина

18
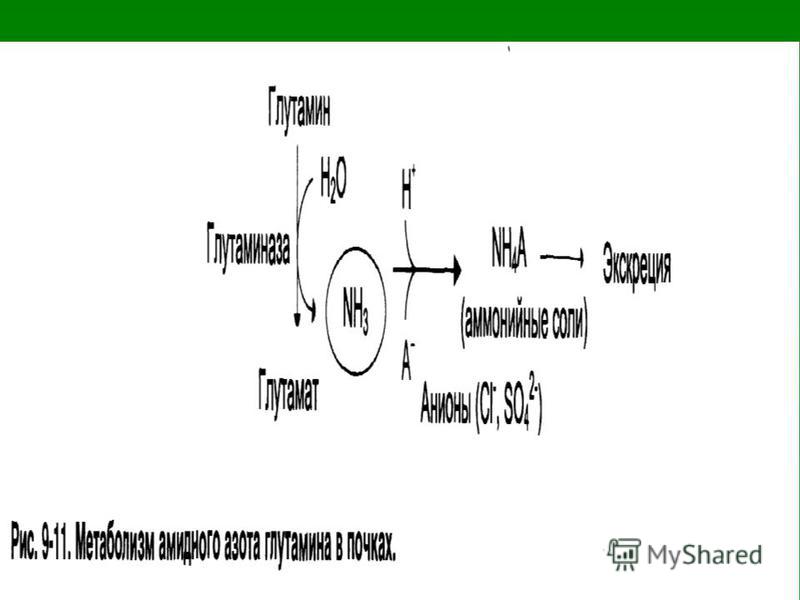
Глутаминаза

21
Синтез аспарагина

22
МОЧЕВИНА МОЧЕВИНА У человека инактивация аммиака осуществляется прежде всего за счет синтеза мочевины, часть NH 3 выводится непосредственно почками. У человека инактивация аммиака осуществляется прежде всего за счет синтеза мочевины, часть NH 3 выводится непосредственно почками.

23
АММОНИОТЕЛИЧЕСКИЕ ОРГАНИЗМЫ У разных видов позвоночных инактивация и выведение аммиака производятся различными способами. Живущие в воде животные выделяют аммиак непосредственно а воду; например, у рыб он выводится через жабры (аммониотелические организмы). У разных видов позвоночных инактивация и выведение аммиака производятся различными способами. Живущие в воде животные выделяют аммиак непосредственно а воду; например, у рыб он выводится через жабры (аммониотелические организмы).

24
УРЕОТЕЛИЧЕСКИЕ ОРГАНИЗМЫ Наземные позвоночные, в том числе человек, выделяют лишь небольшое количество аммиака, а основная его часть превращается в мочевину (уреотелические организмы). Наземные позвоночные, в том числе человек, выделяют лишь небольшое количество аммиака, а основная его часть превращается в мочевину (уреотелические организмы).
. Наземные позвоночные, в том числе человек, выделяют лишь небольшо")
25
УРИКОТЕЛИЧЕСКИЕ ОРГАНИЗМЫ Птицы и рептилии, напротив, образуют мочевую кислоту, которая в связи с экономией воды выделяется преимущественно в твердом виде (урикотелические организмы). Птицы и рептилии, напротив, образуют мочевую кислоту, которая в связи с экономией воды выделяется преимущественно в твердом виде (урикотелические организмы).
. Птицы и рептилии, напротив, образуют мочевую кислоту, которая в св")
26
Синтез мочевины Мочевина в противоположность аммиаку это нейтральное и нетоксичное соединение. Небольшая молекула мочевины может проходить через мембраны, а также из-за ее хорошей растворимости в воде мочевина легко переносится кровью и выводится с мочой. Мочевина в противоположность аммиаку это нейтральное и нетоксичное соединение. Небольшая молекула мочевины может проходить через мембраны, а также из-за ее хорошей растворимости в воде мочевина легко переносится кровью и выводится с мочой.

27
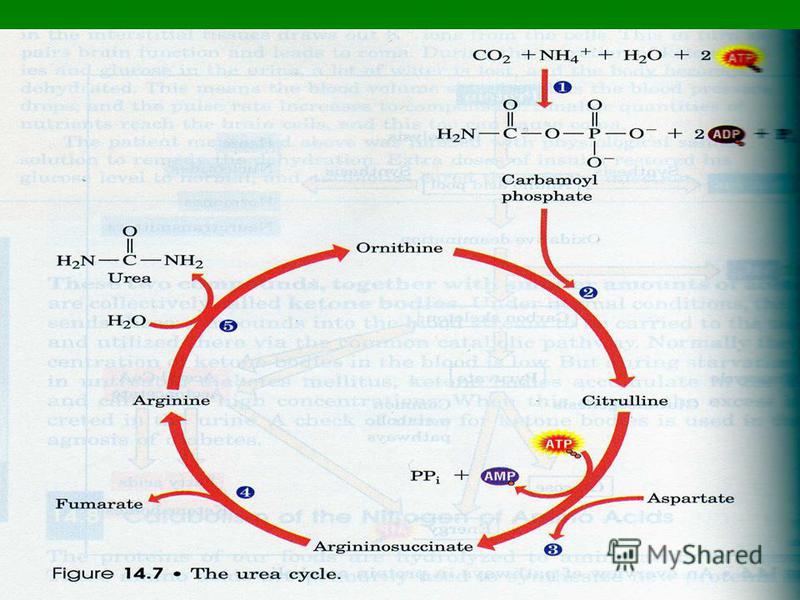
СТАДИИ СИНТЕЗА МОЧЕВИНЫ Мочевина образуется в результате циклической последовательности реакций, протекающих в печени. Мочевина образуется в результате циклической последовательности реакций, протекающих в печени. Оба атома азота берутся из свободного аммиака и за счет дезаминирования аспартата, карбонильная группа из гидрокарбоната. Оба атома азота берутся из свободного аммиака и за счет дезаминирования аспартата, карбонильная группа из гидрокарбоната.

28
Первая реакция На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карбамилфосфат. На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карбамилфосфат.
![Первая реакция На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карбамилфосфат. На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карба](http://images.myshared.ru/17/1098910/slide_28.jpg "Первая реакция На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карбамилфосфат. На первой стадии, реакция [1], из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется карба")
29
Синтез карбамилфосфата

30
Вторая стадия Вторая стадия На следующей стадии, реакция [2], карбамоильный остаток переносится на орнитин с образованием цитруллина. Для этой реакции вновь необходима энергия в форме АТФ, который при этом расщепляется на АМФ и дифосфат. На следующей стадии, реакция [2], карбамоильный остаток переносится на орнитин с образованием цитруллина. Для этой реакции вновь необходима энергия в форме АТФ, который при этом расщепляется на АМФ и дифосфат.
![Вторая стадия Вторая стадия На следующей стадии, реакция [2], карбамоильный остаток переносится на орнитин с образованием цитруллина. Для этой реакции вновь необходима энергия в форме АТФ, который при этом расщепляется на АМФ и дифосфат. На следующей](http://images.myshared.ru/17/1098910/slide_30.jpg "Вторая стадия Вторая стадия На следующей стадии, реакция [2], карбамоильный остаток переносится на орнитин с образованием цитруллина. Для этой реакции вновь необходима энергия в форме АТФ, который при этом расщепляется на АМФ и дифосфат. На следующей")
31
Синтез цитруллина

32
Третья стадия Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3]. Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3].
![Третья стадия Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3]. Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3].](http://images.myshared.ru/17/1098910/slide_32.jpg "Третья стадия Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3]. Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином [3].")
33
Синтез аргининосукцината

34
СТАДИИ СИНТЕЗА МОЧЕВИНЫ Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от аргининосукцината приводит к аргинину [4],
![СТАДИИ СИНТЕЗА МОЧЕВИНЫ Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от аргининосукцината приводит к аргинину [4],](http://images.myshared.ru/17/1098910/slide_34.jpg "СТАДИИ СИНТЕЗА МОЧЕВИНЫ Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от аргининосукцината приводит к аргинину [4],")
35
Четвертая стадия

36
Пятая стадия из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины. из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины.
![Пятая стадия из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины. из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины.](http://images.myshared.ru/17/1098910/slide_36.jpg "Пятая стадия из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины. из которого в результате гидролиза образуется мочевина [5]. Остающийся орнитин вновь включается в цикл мочевины.")
37
Пятая стадия

41
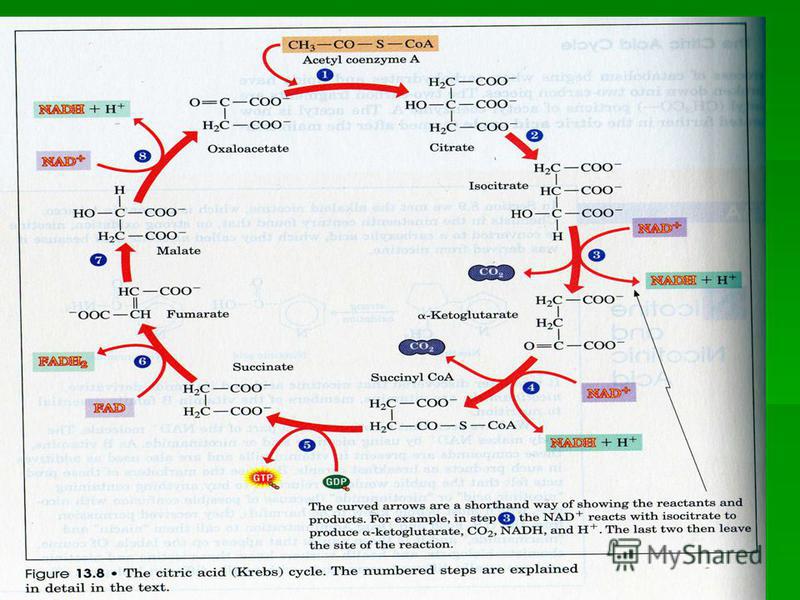
ВЕЛОСИПЕД КРЕБСА Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7] через малат переходить в оксалоацетат, который за счет трансаминирования [9] далее прекращается в аспартат. Последний также вновь вовлекается в цикл мочевины. Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7] через малат переходить в оксалоацетат, который за счет трансаминирования [9] далее прекращается в аспартат. Последний также вновь вовлекается в цикл мочевины.
![ВЕЛОСИПЕД КРЕБСА Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7] через малат переходить в оксалоацетат, который за счет трансаминирования [9] далее прекращается в аспартат. Последний также вновь вовлекае](http://images.myshared.ru/17/1098910/slide_41.jpg "ВЕЛОСИПЕД КРЕБСА Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7] через малат переходить в оксалоацетат, который за счет трансаминирования [9] далее прекращается в аспартат. Последний также вновь вовлекае")
42
Регенерация аспартата

44
Взаимосвязь цикла мочевинообразования и ЦТК

45
ЭНЕРГОЗАВИСИМЫЙ ПРОЦЕСС Биосинтез мочевины требует больших затрат энергии. Энергия поставляется за счет расщепления четырех высокоэнергетических связей: двух при синтезе карбамилфосфата и двух (!) при образовании аргининосукцината (АТФ АМФ + PPi, РРi 2Pi). Биосинтез мочевины требует больших затрат энергии. Энергия поставляется за счет расщепления четырех высокоэнергетических связей: двух при синтезе карбамилфосфата и двух (!) при образовании аргининосукцината (АТФ АМФ + PPi, РРi 2Pi).
 при образовании аргининосукцината (АТФ АМФ + PPi, РРi")
46
КОМПАРТМЕНТАЛИЗАЦИЯ Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента: митохондрии и цитоплазму. Прохождение через мембрану промежуточных соединений цитруллина и орнитина возможно только с помощью переносчиков. Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента: митохондрии и цитоплазму. Прохождение через мембрану промежуточных соединений цитруллина и орнитина возможно только с помощью переносчиков.

47
АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ Скорость синтеза мочевины определяется первой реакцией цикла [1]. Карбамоилфосфатсинтаза активна только в присутствии N- ацетилглутамата. Состояние обмена веществ (уровень аргинина, энергоснабжение) сильно зависит от концентрации этого аллостерического эффектора. Скорость синтеза мочевины определяется первой реакцией цикла [1]. Карбамоилфосфатсинтаза активна только в присутствии N- ацетилглутамата. Состояние обмена веществ (уровень аргинина, энергоснабжение) сильно зависит от концентрации этого аллостерического эффектора. Скорость синтеза мочевины определяется первой Скорость синтеза мочевины определяется первой
![АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ Скорость синтеза мочевины определяется первой реакцией цикла [1]. Карбамоилфосфатсинтаза активна только в присутствии N- ацетилглутамата. Состояние обмена веществ (уровень аргинина, энергоснабжение) сильно з](http://images.myshared.ru/17/1098910/slide_47.jpg "АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ Скорость синтеза мочевины определяется первой реакцией цикла [1]. Карбамоилфосфатсинтаза активна только в присутствии N- ацетилглутамата. Состояние обмена веществ (уровень аргинина, энергоснабжение) сильно з")
48
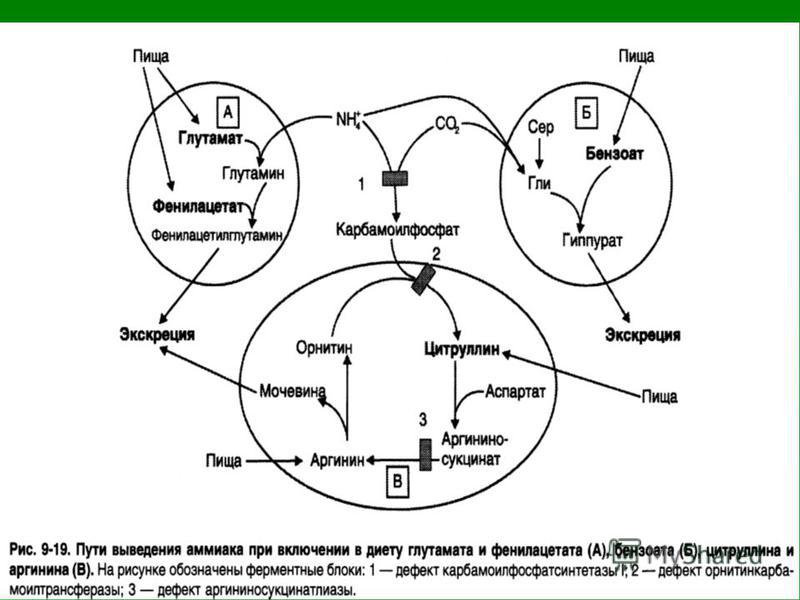
Наследственные нарушения орнитинового цикла и их симптомы

50
Включение безазотистого остатка АК в ЦТК

51
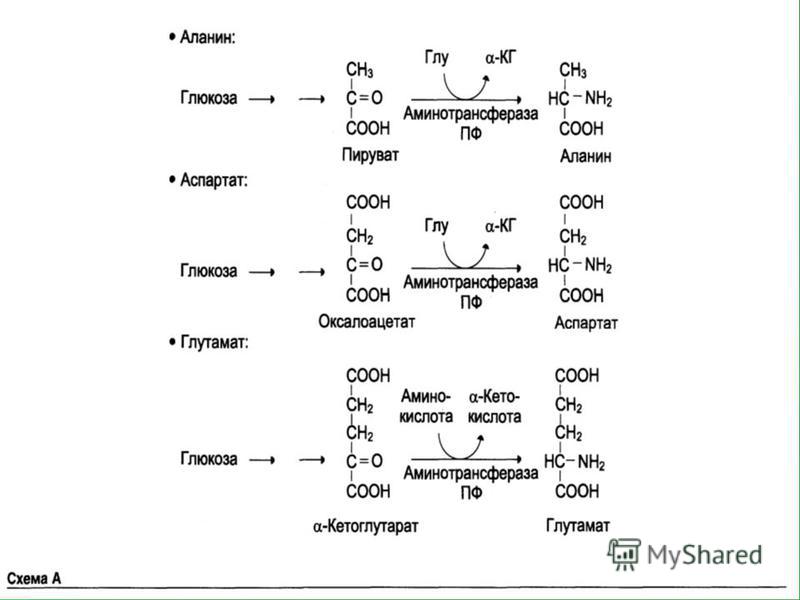
Пути биосинтеза заменимых АК

53
Глюкозо-аланиновый цикл

55
Обмен аммиака

Еще похожие презентации в нашем архиве:







 по теме: обмен веществ и энергии в клетке.")


 и синтез глюкозы (глюконеогенез) Автор – доцент кафедры биохимии Рыскина Е.А.")




 (обмен веществ) полный процесс превращения химических веществ в организме, обеспечивающих его.")



 – совокупность протекающих в клетках химических превращений, обеспечивающих их рост, жизнедеятельность,")


