Скачать презентацию
Идет загрузка презентации. Пожалуйста, подождите

1
МАТЕМАТИЧЕСКОЕ моделирование в естествознании Математическое моделирование практически единственный метод решения научных задач особой сложности. Метод сводится к построению математической модели, позволяющей представить систему или процесс, и проведению экспериментов на модели.

2
Среда Организм Аутэкология – организм во взаимодействии со средой как система, отражающая видовые особенности Экологическая физиология – функциональные взаимодействия организма и среды Факториальная экология – воздействие факторов среды на организмы.

3
Окружающая среда Живая система Абиотическ ая система

4
Зоогеографическое правило Бергмана: Среди близких форм гомойотермных животных те, что обитают в более холодном климате, как правило имеют более крупные размеры. Причиной этого является то, что теплопродукция имеет более высокий аллометрический коэффициент зависимости от массы тела, чем теплоотдача (первый имеет значение около 0,75, а второй – около 0,5). Поэтому поддерживать постоянную температуру тела крупным легче, чем мелким.

5
S 1 = 6 a 2 S 2 = 6 (2a) 2 = 24 a 2 V 1 = a 3 V 2 = (2a) 3 = 8 a 3 T 1 = V 1 / S 1 = a 3 / 6 a 2 = a / 6 T 2 = V 2 / S 2 = 8a 3 / 24 a 2 = a / 3 Математическая модель. Представим себе двух животных, имеющих тело в виде правильных кубов со сторонами а у первого и 2 а - у второго животного. Таким образом, отношение V / S (фактически - отношение теплопродукции к теплоотдаче!) у второго животного в два раза выгоднее для условий, где теплопродукция должна быть больше, чем теплоотдача (т.е. для холодного климата).
 2 = 24 a 2 V 1 = a 3 V 2 = (2a) 3 = 8 a 3 T 1 = V 1 / S 1 = a 3 / 6 a 2 = a / 6 T 2 = V 2 / S 2 = 8a 3 / 24 a 2 = a / 3 Математическая модель. Представим себе двух животных, имеющих тело в виде правильных кубов со сторонами а")
6
Правило Аллена: У гомойотермных животных выступающие части (теплоотдатчики – в данном примере - уши), придающие более грациозный облик, больше у тех, кто живет в более жарком климате. Песец (Арктика) Красная лисица (Умеренная зона) Фенек (Северная Африка) В определенном смысле правило Аллена (в отличие правила Бергмана) объясняет не стремление избежать переохлаждения, А стремление избежать перегрева.
, придающие более грациозный облик, больше у тех, кто живет в более жарком климате. Песец (Арктика) Красная лисица (Умеренная зона) Фенек (Северная Аф")
7
S 1 =... V 1 =... T 1 = V 1 / S 1 =... S 2 =... V 2 =... T 2 = V 2 / S 2 =... Математическая модель. Представим себе двух животных, имеющих тело, образованное правильными кубиками со стороной а. Подсчитаем, сколько (в процентах) тепла сэкономит первое животное по сравнению со вторым: T % = (Т 2 - Т 1 )/ Т 2 * 100% =... (
 тепла сэкономит перво")
8
Арктический вид Температура тела 37°С Средняя температура среды 0°С Европейский вид Температура тела 37°С Средняя температура среды 12°С Африканский вид Температура тела 37°С Средняя температура среды 25°С Температура тела 37°С Средняя температура среды 25°С Различия в длине ушей у трех видов лисиц, обитающих в разных географических областях (по Н. Грину и др., 1993)

10
Задание 1

11
Задание 2

12
«Сопоставимый» энергетический обмен (А.И. Зотин, А.А. Зотин, 1999) -это коэффициент a в аллометрическом уравнении зависимости интенсивности обмена (R)от характерных для вида размеров (массы M)): R = a M b Биологический смысл этого коэффициента - ожидаемая величина энергозатрат при единичной массе тела. Коэффициент b - (аллометрический экспонент) мало изменяется у разных видов. Проф. А.И. Зотиным было показано, что величина a может служить показателем организованности, упорядоченности (т.е. сложности), как выражение термодинамического потенциала (диссипативной функции по И. Р. Пригожину).
 -это коэффициент a в аллометрическом уравнении зависимости интенсивности обмена (R)от характерных для вида размеров (массы M)): R = a M b Биологический смысл этого коэффициента - ожид")
13
Гипотеза «термальных барьеров» (Зотин, Зотин, 1999). Согласно этой гипотезе, пойкилотермные (эндотермные) животные не могут иметь величину сопоставимого обмена более (5-8 м Вт/г), иначе они погибнут от перегрева. Это - первый тепловой барьер, достигнутый у прямокрылых и однопроходных млекопитающих. Следование тенденции прогрессивной эволюции привело к развитию гомойотермии. Однако по мере биоэнергетического прогресса, т.е. увеличения a, был достигнут второй тепловой барьер в м Вт/г (у воробьиных птиц). Первый тепловой барьер Второй тепловой барьер
. Согласно этой гипотезе, пойкилотермные (эндотермные) животные не могут иметь величину сопоставимого обмена более (5-8 м Вт/г), иначе они погибнут от перегрева. Это - первый тепловой барьер, достигн")
14
ПРОДОЛЖЕНИЕ ТЕНДЕНЦИИ К ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ БЫЛО МОЖНО ТОЛЬКО ПРИ ПЕРЕХОДЕ К НОВОМУ (НЕБИОХИМИЧЕСКОМУ) СПОСОБУ ПОЛУЧЕНИЯ И РАСХОДОВАНИЯ ЭНЕРГИИ (НЕ ИЗ ПИЩИ) И СВЯЗАННЫМ С ЭТИМИ ПРОЦЕССАМИ ИСПОЛЬЗОВАНИЕМ НОВЫХ РЕСУРСОВ). Это стало возможным с появлением цивилизации, которая позволила источники энергии вне организма. Такая способность возможна только развитых системах получения, хранения и использование информации (сознание), но и одновременно с социализацией - (появлением общества)
 СПОСОБУ ПОЛУЧЕНИЯ И РАСХОДОВАНИЯ ЭНЕРГИИ (НЕ ИЗ ПИЩИ) И СВЯЗАННЫМ С ЭТИМИ ПРОЦЕССАМИ ИСПОЛЬЗОВАНИЕМ НОВЫХ РЕСУРСОВ). Это стало возможным с появл")
15
Владимир Иванович Вернадский ( ) Такая тенденция согласуется с представлениями В.И. Вернадского о ускорении круговорота веществ и усиления потока энергии по мере эволюции биосферы.
 Такая тенденция согласуется с представлениями В.И. Вернадского о ускорении круговорота веществ и усиления потока энергии по мере эволюции биосферы.")
17
Потребление энергии на одного человека в сутки (к Дж). Лет назад
. Лет назад")
18
Стадии исторического развития человечества и значения коэффициента a (м Вт/г)
")
20
Смена экологических функций сопровождалась быстрым ростом численности человечества.

21
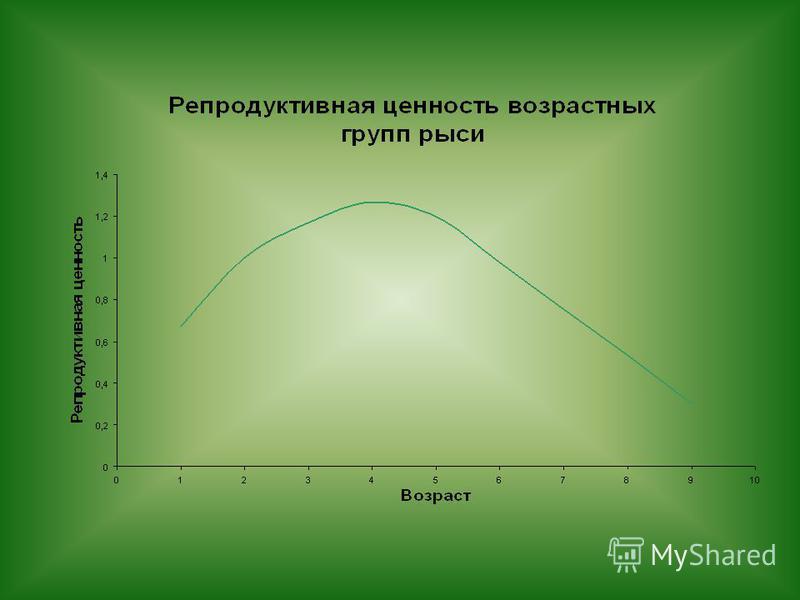
Эти характеристики служат для построения демографических таблиц, по параметрам которых можно рассчитать приспособленность или (чистую скорость размножения R 0 ), ожидаемую продолжительность жизни в возрасте X (E X ), показывающий, сколько еще осталось в среднем прожить, а также «репродуктивную ценность» в возрасте X (V X ), показывающий, сколько в среднем принесет потомков особь этого возраста за оставшуюся жизнь. Математическое моделирование
, ожидаемую продолжительность жизни в возрасте X (E X ), показывающий, сколько еще осталос")
22
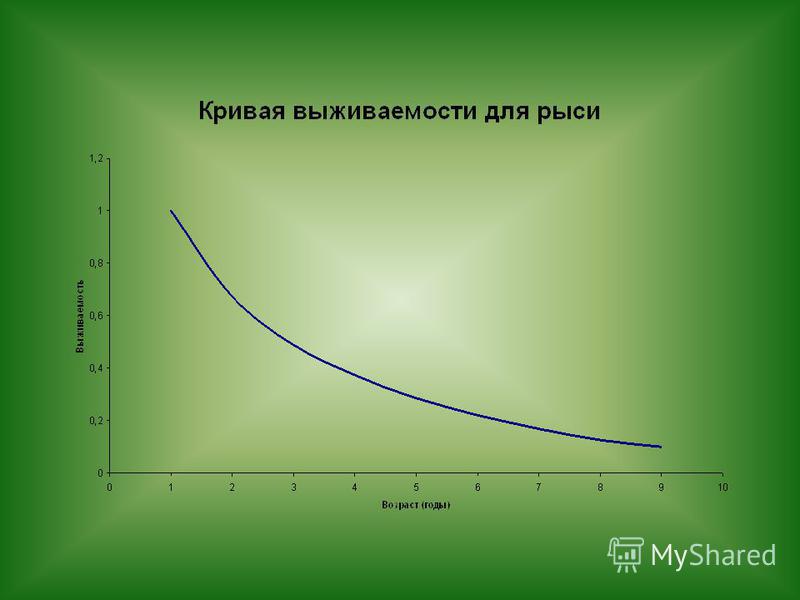
Для примера приведем таблицу для рыси (Lynx lynx) в 70-е-80-е годы XX в., когда наблюдался спад численности этого хищника на Среднем Урале.
 в 70-е-80-е годы XX в., когда наблюдался спад численности этого хищника на Среднем Урале.")
26
Чистая скорость размножения связана с мальтузианским параметром r, а все параметры связывает так называемое основное демографическое уравнение Эйлера-Лотки:

27
МАЛЬТУЗИАНСКИЙ ПАРАМЕТР, так иногда называют врожденную (специфич.) скорость естественного роста популяции (по имени Т. Мальтуса). Обозначается символом г. (см. Уравнения Ферхульста-Пирла). Эта величина является мерой мгновенной удельной скорости изменения размера популяции и выражается как число особей на единицу времени и на одну особь, т. е. имеет размерность 1/время. В замкнутой популяции М. п. определяется как мгновенная удельная рождаемость b минус мгновенная удельная смертность d. В незамкнутой популяции: r = (рождаемость + иммиграция) (смертность + + эмиграция). Если удельная рождаемость превышает удельную смертность (b>d), то популяция растет и r положительна; если смертность превышает рождаемость (b скорость естественного роста популяции (по имени Т. Мальтуса). Обозначается символом г. (см. Уравнения Ферхульста-Пирла). Эта величина является мерой мгновенной удельной скорости изм")
28
Одной и той же приспособленности можно достичь, либо интенсивно размножаясь, но имея большую смертность, либо «вкладывать» в доведение всего (немногочисленного) потомства до репродуктивного возраста.
 потомства до репродуктивного возраста.")
29
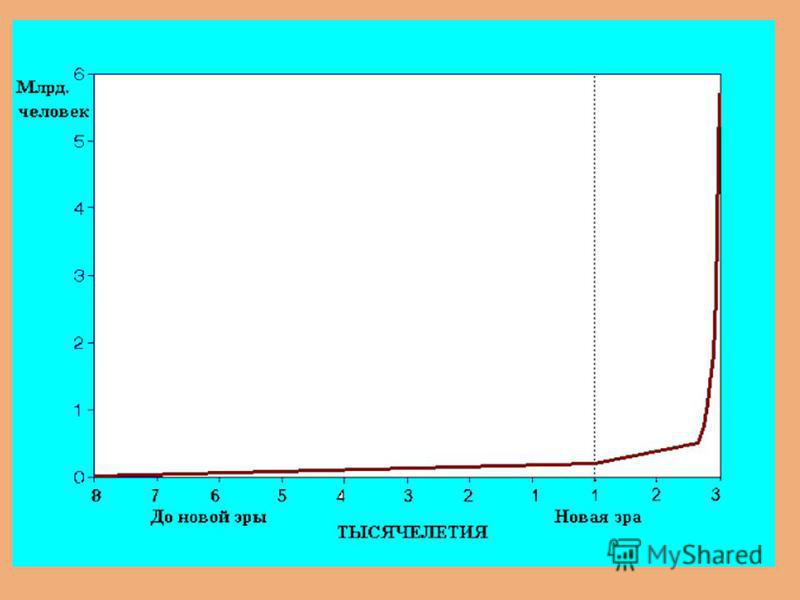
Динамика численности человечества Демографический взрыв и демографический переход Математическое моделирование.

33
Моделирование роста человечества Сергей Петрович Капица

35
Verhulst, P.-F. "Recherches mathematiques sur la loi d'accroissement de la population." Nouv. mem. de l'Academie Royale des Sci. et Belles-Lettres de Bruxelles 18, 1-41, 1845.

36
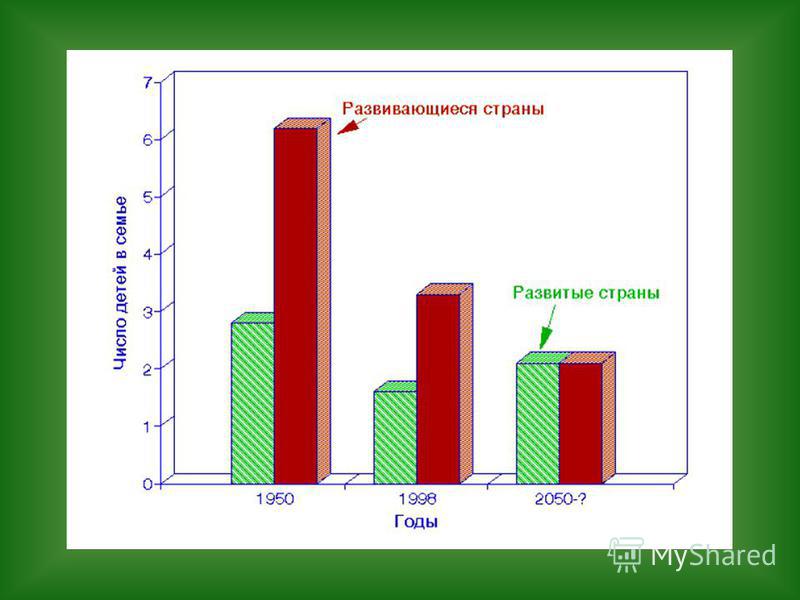
« Демографический взрыв» и «демографический переход» Рост численности человечества носил гиперболический характер, особенно после «промышленной революции» Деннис Медоуз Одновременно (особенно в последнее время) наблюдается «демографический переход» (более заметный в «развитых странах»- некоторое снижение темпов роста одновременно с постарением населения из-за синхронного снижения и рождаемости и смертности
 наблюдается «демографический переход» (")
37
Современный этап: достоинства и вызовы «Демографический переход» - признак стабилизации?

39
Незаметное сначала истощение медленно возобновляющихся ресурсов (с повышением их относительного потребления) должно смениться резким падением их доступного количества Истощение ресурсов - возможная причина спада
 должно смениться резким падением их доступного количества Истощение ресурсов - возможная причина спада")
40
N = rN(1-N/K) t Роберт Мэй May R.M. Biological populations obeing difference equations: stable points, cycles, snd chaos. - J. Theor. Biol., 1975, vol.51, N 2, p
 t Роберт Мэй May R.M. Biological populations obeing difference equations: stable points, cycles, snd chaos. - J. Theor. Biol., 1975, vol.51, N 2, p. 511-524")
41
«Технократический оптимизм» вызывает сомнения Следовало бы обратить больше внимания на общность человека с другими биологическими видами, учитывая багаж, накопленный классической экологией

Еще похожие презентации в нашем архиве:










 137 ауд. 8 к.")


 по теме: Популяции")












